영장류 목차 진화의 역사 구별되는 특징 해부학, 생리학 그리고 형태학 습성 서식지 및 분포 사람과의 상호 작용 함께 읽기 각주 외부 링크 둘러보기 메뉴《Mammal Species of the World: A Taxonomic and Geographic Reference》62265494210908710.1007/BF02099995“영장류(브리태니커 백과사전 온라인)”“초행렬 접근을 이용한 영장류 계통발생 및 분기 싯점 추산(Estimating the phylogeny and divergence times of primates using a supermatrix approach)”27747001986089110.1186/1471-2148-9-259“Molecular and Genomic Data Identify the Closest Living Relative of Primates”10.1126/science.1147555“유인원 기원에 관한 새로운 관점(New perspectives on anthropoid origins)”2010PNAS..107.4797W28419172021210410.1073/pnas.0908320107“Nova – Meet Your Ancestors”“플레시아다피스(Plesiadapis)”1047378010.1007/PL00006562“Scientists Push Back Primate Origins From 65 Million To 85 Million Years Ago”1196155210.1038/416726a“Molecular Remodeling of Members of the Relaxin Family During Primate Evolution”11230540“Development and Application of a Phylogenomic Toolkit: Resolving the Evolutionary History of Madagascar's Lemurs”22591131824577010.1101/gr.7265208원본 문서10.1007/s10764-008-9317-y“Primate Evolution”보존된 문서“Complete Primate Skeleton from the Middle Eocene of Messel in Germany: Morphology and Paleobiology”26835731949208410.1371/journal.pone.0005723“Evolutionary Biology of Tarsiers”“Rapid electrostatic evolution at the binding site for cytochrome c on cytochrome c oxidase in anthropoid primates”10883651585167110.1073/pnas.0409714102“Anthropoid primates from the Oligocene of Pakistan (Bugti Hills): Data on early anthropoid evolution and biogeography”11508601593710310.1073/pnas.0503469102“Timing the Origin of New World Monkeys”1283265310.1093/molbev/msg1721042326810.1002/(SICI)1096-8644(199908)109:4<541::AID-AJPA9>3.0.CO;2-N1785578610.1159/00010514810.1002/ajp.1350080307495893310.1126/science.154.3756.155910.1007/BF0237382710.1023/A:102636730747010.1006/jhev.1998.0263“Archonta: Primates”원본 문서“"Primates" (On-line)”“Cone topography and spectral sensitivity in two potentially trichromatic marsupials, the quokka (Setonix brachyurus) and quenda (Isoodon obesulus)”15998611588841110.1098/rspb.2004.3009“Photosensitive and photostable pigments in the retinae of Old World monkeys”0022-0949205112710.1016/S0169-5347(03)00012-01468653810.1554/03-16810.1016/S0169-5347(03)00012-078552410.1086/40931010.1086/28438510.1007/BF0165914881003410.1002/ajpa.133043020710.2307/240879010.1007/BF026937401178699010.1002/ajpa.1001110.1017/S09528369020008571138088410.1046/j.1365-294X.2001.01262.x215765110.1159/000156428964278810.1037/0735-7036.112.2.2071245922010.1016/S0028-3932(02)00152-5“Recognizing facial cues: Individual discrimination by chimpanzees (Pan troglodytes) and rhesus monkeys (Macaca mulatta)”20187441073931110.1037/0735-7036.114.1.471038511410.1038/2134510.1007/BF023816121145916310.1037/0735-7036.115.2.159964278210.1037/0735-7036.112.2.11910.1037/0096-3445.130.3.505“Relaxed selective pressure on an essential component of pheromone transduction in primate evolution”1522921263169810.1073/pnas.0636123100〈Nonhuman Primate Communication〉10.1159/000156551“Theropithecus gelada”“Geographic and climatic control of primate diversity”41248764450610.1073/pnas.92.17.787410.1007/BF02382060“Zoonoses Acquired from Pet Primates”“Declaration on Great Apes”원본 문서“Spanish parliament approves 'human rights' for apes”“Of great apes and men”“The HSUS Applauds the U.S. Senate for Unanimously Passing the Captive Primate Safety Act”원본 문서10.1525/aa.2004.106.2.346“IPPL News: The US Pet Monkey Trade”원본 문서“The Beginnings of Research in Space Biology at the Air Force Missile Development Center, 1946-1952”“Monkeys as Helpers To Quadriplegics At Home”“The supply and use of primates in the EU”원본 문서“Genomic divergences between humans and other hominoids and the effective population size of the common ancestor of humans and chimpanzees”12352771117089210.1086/318206“A Demographic Analysis of Primate Research in the United States”원본 문서《Statistics of scientific procedures on living animals: Great Britain 2005》“Nonhuman Primates: Research Animals”원본 문서“Directive 86/609”원본 문서10.1002/1520-6505(2001)10:1<16::AID-EVAN1010>3.0.CO;2-O“Status, conservation and management of primates in India”원본 문서10.1046/j.1365-3008.2001.00150.x“Predicting extinction risk in declining species”16907721107570610.1098/rspb.2000.123410.1046/j.1523-1739.1995.9051107.x10.1080/09670879722902210.1525/aa.2002.104.4.1184〈Primate〉“Primates in Northeast India: an overview of their distribution and conservation status”원본 문서10.1023/B:IJOP.0000043348.06255.7f“Conservation of the Javan gibbon Hylobates moloch: population estimates, local extinction, and conservation priorities”10.1023/B:IJOP.0000019152.83883.1c10.1111/j.1600-0587.1994.tb00098.x10.2307/2404743“Primates in Peril: The World's 25 Most Endangered Primates, 2006 – 2008”10.1896/052.022.0101원본 문서10.1046/j.1523-1739.2000.99230.x10.1007/s10764-005-4368-9영장류영장류 정보넷(Primate Info Net)동물다양성웹(Animal Diversity Web)의 영장류(Primates) 정보교토 대학의 영장류 연구소고해상도 세포구축학적 영장류 뇌 사진/도해(High-Resolution Cytoarchitectural Primate Brain Atlases)유러피안 영장류 네트워크(EUPRIM-Net: European Primate Network)영장류 사진: 자연사 콜렉션(PrimateImages: Natural History Collection)생물의 나무(Tree of Life) 웹프로젝트(web project)의 영장류ehehsh851066684047256-500569360
영장류칼 폰 린네가 명명한 분류군
포유류원원류여우원숭이로리스갈라고안경원숭이진원류원숭이유인원인간아메리카아프리카아시아베르트부인쥐여우원숭이산악고릴라백악기팔레오세플레시아다피스원원류진원류마다가스카르여우원숭이로리스안경원숭이원숭이유인원분류학자곡비원아목직비원아목광비원소목협비원소목꼬리감는원숭이고함원숭이다람쥐원숭이구세계원숭이개코원숭이마카크긴팔원숭이대형 유인원쥐목토끼목나무두더지목날원숭이목플레시아다피스목영장목영장상목포유강진수하강날원숭이류나무두더쥐류영장동물설치동물설치류토끼류플레시아다피스분지학단계통군곡비원아목여우원숭이로리스과갈라고과족제비여우원숭이과여우원숭이과갈라고과로리스과에오세아디피드오모미드마다가스카르뗏목 사건아이아이아이아이과여우원숭이하목직비원아목안경원숭이과안경원숭이하목원원류원숭이하목신세계원숭이광비원소목구세계원숭이사람유인원협비원소목에오시미드에오세대륙 이동설마이오세베마라하양털여우원숭이여우원숭이황금궁전티티원숭이린네자연의 체계사람속시미아여우원숭이속박쥐속세발가락나무늘보속안드로포모르파앙리 드 블랭빌ChiropteraInsectivoraCarnivora원원류진원류원원류안경원숭이진원류잡종잡종 지역동물원겔라다속망토개코원숭이거미원숭이목도리여우원숭이양안시뼈가 융기된곡비원류후안와봉직비원류안드로포이드cm3감각 인지운동 명령의식적인 사유언어신피질대뇌 피질후각촉각시각주머니쥐쇄골팔이음뼈어깨 관절어깨뼈흉각고함원숭이거미원숭이양털원숭이거미원숭이과주둥이치열참빗진수하강포유류3색성 색각중생대망막4색성 색각유대류단색성 색각올빼미원숭이부시베이비옵신유전자 복제X 염색체대립 유전자유전자 복제진원류성 결정자연 선택수렴 진화부시베이비인드리시파카아바히인드리족제비여우원숭이안경원숭이긴팔원숭이무리키거미원숭이양털원숭이오랑우탄침팬지고릴라오스트랄로피테신초기 인류리처드 랭햄사회 조직고함원숭이곰비 천 국립공원제인 구달망토개코원숭이거미원숭이보노보겔라다사회적 그루밍피부의 기생충타이 숲다이애나원숭이캠벨모나원숭이작은흰코원숭이서부붉은콜로부스왕콜로부스검댕망가베이침팬지붉은꼬리원숭이서부붉은콜로부스푸른원숭이울프모나원숭이망토게라자검은볏망가베이앨런습지원숭이다람쥐원숭이카푸친원숭이여우원숭이로리스안경원숭이신세계원숭이페로몬서골비기관탄수화물지질아미노산비타민미네랄고함원숭이흑백콜로부스족제비여우원숭이마모셋앞니아이아이회색뺨망가베이겔라다안경원숭이카푸친원숭이무척추동물도마뱀다람쥐박쥐척추동물침팬지서부붉은콜로부스열대 우림생태학개코원숭이파타스원숭이겔라다사람우림망그로브일본원숭이혼슈 섬바르바리마카크아틀라스 산맥검은들창코원숭이산악고릴라코주부원숭이브라자원숭이알렌습지원숭이붉은털원숭이회색랑구르온대 기후사람헤르페스 바이러스헤르페스 B 바이러스천연두 바이러스홍역에볼라광견병마르부크 바이러스바이러스성 감염개인국제 연합세계 인권 선언애완동물모델 생물우주 비행서비스 동물꼬리감는원숭이동물원유전적 다양성신경과학히말라야원숭이마카크푸른원숭이침팬지개코원숭이다람쥐원숭이마모셋유럽 연합국제 자연 보호 연맹애완동물팜유수마트라오랑우탄애완동물게논드릴원숭이마다가스카르애완동물모델 생물히말라야원숭이세계에서 멸종 위험이 가장 높은 25종의 영장류비단시파카델라쿠르랑구르흰머리랑구르회색정강이두크통킹들창코랑구르하이난검은볏긴팔원숭이크로스강고릴라수마트라오랑우탄왈드론붉은콜로부스
(function()var node=document.getElementById("mw-dismissablenotice-anonplace");if(node)node.outerHTML="u003Cdiv class="mw-dismissable-notice"u003Eu003Cdiv class="mw-dismissable-notice-close"u003E[u003Ca tabindex="0" role="button"u003E숨기기u003C/au003E]u003C/divu003Eu003Cdiv class="mw-dismissable-notice-body"u003Eu003Cdiv id="localNotice" lang="ko" dir="ltr"u003Eu003Cpu003Eu003Ca href="/wiki/%EC%9C%84%ED%82%A4%EB%B0%B1%EA%B3%BC:%EA%B3%BC%ED%95%99%EC%9D%98_%EB%8B%AC_%EC%97%90%EB%94%94%ED%84%B0%ED%86%A4" title="위키백과:과학의 달 에디터톤"u003E과학의 달 에디터톤u003C/au003E이 4월 30일까지 진행됩니다.nu003C/pu003Eu003C/divu003Eu003C/divu003Eu003C/divu003E";());
영장류
둘러보기로 가기
검색하러 가기
 올리브개코원숭이 (Papio anubis) | ||||||||||||
생물 분류 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||
아목 | ||||||||||||
| ||||||||||||
 사람을 제외한 영장류의 분포 지역 | ||||||||||||
영장류[1](靈長類)는 생물분류학적으로 영장목(靈長目)에 속하는 포유류로 원원류(여우원숭이와 로리스, 갈라고, 안경원숭이 포함)와 진원류(원숭이와 유인원)를 포함하고 있으며 인간도 영장류에 속한다. [2] 전 세계에 살고 있는 사람을 제외하면, 대부분의 영장류는 아메리카와 아프리카 그리고 아시아의 열대 또는 아열대 지역에 살고 있다.[3] 크기 면에서 영장류는 30g의 베르트부인쥐여우원숭이부터 200kg의 산악고릴라까지 있다. 화석 기록에 의하면, 영장류의 원시 조상은 약 6,500만년 전의 백악기 후기까지 거슬러 올라가며, 알려진 영장류 중 가장 오랜 영장류는 약 5,500 ~ 5,800만년 전의 팔레오세 후기의 플레시아다피스이다.[4] 분자 시계를 통한 연구는 약 8,500만년 전 경의 백악기-중기로부터 기원하는 선조들보다 더 오래전에 영장류가 분리되었음을 시사하고 있다.[4]
영장목은 전통적으로 크게 원원류와 진원류라는 두 그룹으로 나누어 왔다. 원원류는 초기 영장류의 특징을 좀더 많이 지니고 있으며, 마다가스카르의 여우원숭이, 로리스 그리고 안경원숭이를 포함하고 있다. 진원류는 원숭이와 유인원을 포함하고 있다. 좀더 최근의 분류학자들은 영장목을 안경원숭이를 제외한 원원류로 이루어진 곡비원아목(曲鼻猿亞目, ‘구부러진 코를 지닌 영장류’)과 안경원숭이와 진원류로 이루어진 직비원아목(直鼻猿亞目, ‘코가 마르는 영장류’)으로 나누고 있다. 진원류 또한 크게 두 그룹으로 나눈다. 남아메리카와 중앙아메리카에 사는 신세계원숭이 또는 광비원소목(廣鼻猿小目, ‘코가 넓고 평평한 원숭이류’)과 아프리카와 동남아시아에 사는 구세계원숭이 협비원소목(狹鼻猿小目, ‘코가 좁은 원숭이류’)이다. 신세계원숭이는 꼬리감는원숭이, 고함원숭이, 다람쥐원숭이를 포함하고 있다. 협비원류는 구세계원숭이(개코원숭이와 마카크 등)와 긴팔원숭이 그리고 대형 유인원으로 이루어져 있다. 사람은 아프리카와 남아시아, 동아시아를 벗어나 전 세계에 성공적으로 정착한 현존하는 유일한 협비원류이지만, 화석 기록을 통해 기타 여러 종이 이전에 현재의 유럽에 살았던 것으로 확인된다. 일부 영장류는 2000년대에 발견되었다.
포유류 중에서 가장 지능이 뛰어나고 다양한 능력을 지닌 포유류로 간주되는, 영장류는 광범위한 특징을 보여준다. 일부 영장류(대형 유인원과 개코원숭이를 포함하여)는 나무 위에서 살기 보다는 땅 위에서 주로 살지만, 모든 종들은 나무타기에 적합한 신체 구조를 갖고 있다. 장소를 이동하는 방법으로 나무 사이를 건너뛰는 도약, 두 발 또는 네발로 걷기, 관절을 이용한 걷기, 나뭇가지를 흔들어 양손을 번갈아 매달려 건너가기 등을 사용한다. 영장류는 다른 포유류에 비해 상대적으로 큰 뇌를 갖고 있을 뿐만 아니라, 대부분의 포유류가 지배적인 감각기관으로 갖고 있는 후각을 포기하는 대신, 입체적인 시각 능력이 크게 발달해 있다. 원숭이와 유인원은 이런 특징이 매우 두드러지지만, 로리스원숭이와 여우원숭이는 다소 떨어진다. 일부 영장류는 삼색형 색각이 발달해 있다. 그리고 대부분의 종이 엄지손가락을 다른 손가락과 마주 대할 수 있으며, 일부 종은 물건을 쥘 수 있는 꼬리를 갖고 있다. 많은 종이 암수가 성적이형인데, 이는 암수가 몸무게와 송곳니 크기 그리고 채색 등의 신체적 특징이 서로 다름을 의미한다. 영장류는 비슷한 크기의 다른 포유류보다 성장 속도가 느리고, 뒤늦게 성장하지만, 완전히 자란 후에는 오히려 수명이 더 길다. 일부 종은 혼자 생활하며, 일부는 암수가 짝을 지어 살고, 일부는 수백마리씩 집단적으로 모여 산다.
목차
1 진화의 역사
1.1 진화
1.2 현존하는 영장류 분류
1.3 잡종
2 구별되는 특징
3 해부학, 생리학 그리고 형태학
3.1 성적 이형성
3.2 이동
4 습성
4.1 사회 체계
4.2 이종간 군집
4.3 인지 능력과 의사 소통
4.4 생활사
4.5 먹이
5 서식지 및 분포
6 사람과의 상호 작용
6.1 법적 사회적 상태
6.2 과학 연구에서의 역할
6.3 보존
7 함께 읽기
8 각주
9 외부 링크
진화의 역사
다음은 영장상목의 계통 분류이다.[5][6][7]
|
영장목은 영장상목 계통군의 일부로로, 포유강의 진수하강에 속한다. 최근에 이루어진, 영장류와 날원숭이류 그리고 나무두더쥐류에 대한 분자생물학적 연구에 의하면, 날원숭이류의 두 종은 나무두더쥐류보다는 영장류에 좀더 밀접한 관계가 있다는 것이 밝혀졌으며,[8] 한때 나무두더쥐류를 영장류로 간주하기도 했다.[9] 이들 세 목은 영장동물 계통군을 형성한다. 이 계통군은 설치동물 계통군(설치류와 토끼류로 이루어진)과 합쳐 영장상목 계통군을 형성한다. 영장동물과 영장상목 둘 다 상목으로 분류하기도 한다. 또한 일부 과학자들은 날원숭이류를 영장류의 아목으로 간주하여, 이름에 참 진(眞, true)을 붙여 진영장아목(眞靈長亞目, Euprimates)으로 부르기도 한다.[10]
진화
영장류 계통 분류는 최소 6,500만 전까지 거슬러 올라가는 것으로 추정하고 있지만,[11] 화석 기록으로 남아 있는 가장 오래된 영장류는 백악기 후기의 플레시아다피스(약 5,500~5,800만년 전)이다.[12][13] 분자시계 연구를 포함한 다른 연구를 통해, 영장류 분화의 기원을 백악기 중기 약 8,500만년 전 경으로 추정하고 있다.[14][15][16]
근대 분지학적인 추정에 의하면, 영장목은 단계통군이다. 구부러진 코 또는 젖는 코를 지닌 영장류인, 곡비원아목은 일반적으로 약 6,300만년 전에[17] 원시 영장류 계통에서 분리된 것으로 생각하고 있다.[18] 7개의 곡비원류 과는 5개의 여우원숭이 관련 과와 나머지 로리스과, 갈라고과로 이루어져 있다.[1][19] 이전의 분류 체계는 족제비여우원숭이과를 여우원숭이과에, 갈라고과를 로리스과에 포함시켜 분류하여, 현재의 분류 체계인 5+2개 과 분류 대신에 3+2개 과로 분류를 나누었다.[1]에오세 동안, 북반구 대륙의 대부분은 2개의 그룹 아디피드와 오모미드가 우위를 점하며 살고 있었다.[20][21] 전자는 곡비원류의 일종으로 간주하지만, 현대의 여우원숭이들처럼 촘촘한 치아를 갖고 있지는 않다. 최근의 연구는 이 집단을 다위니우스 마실라에(Darwinius masillae)라는 이름으로 제안하고 있다.[22] 후자는 안경원숭이와 원숭이 그리고 유인윈과 밀접한 관련이 있다. 이 두 그룹이 현존하는 영장류와 정확히 어떻게 관련이 있는지는 명확치 않다. 오모미드는 약 3,000만년 전에 사라졌고,[21] 반면에 아다피드는 약 1,000만년 전까지 살아남았다.[23]

곡비원류 영장류의 일종인 호랑이꼬리여우원숭이
발생학적 연구에 의하면, 마다가스카르의 여우원숭이는 약 7,500만년 전에 로리스하목에서 갈라졌다.[18] 이들 연구는 염색체와 분자적 증거뿐만 아니라 여우원숭이가 다른 곡비원류 영장류보다 서로 밀접한 관련이 있음을 보여준다.[18][24] 그러나 마다가스카르는 1억 6천만년 전에 아프리카 대륙에서, 9천만년 전에 인도에서 분리되었다.[25] 여우원숭이들이 다른 곡비원류 영장류보다 서로 더 밀접한 관계에 있기 때문에, 일단의 매우 적은 규모의 선조 개체군이 5천만년 전과 8천만년 전 사이에 일종의 단일한 뗏목 사건을 통해 마다가스카르로 왔기 때문으로 생각된다.[18][24][25] 아프리카와 인도로부터의 여러 차례의 이주와 같은, 기타 여러 이주 조건들을 시험해봤으나 어느 것도 유전적이며 분자학적인 증거에 의해 뒷받침되지 않았다.[20]
최근까지 아이아이는 곡비원류로 분류하기 어려웠다.[1]아이아이과를 여우원숭이하목 영장류(여우원숭이와 로리스로 분리되는 것보다 더 최근에 여우원숭이 계열에서 분리되는 것을 의미)의 일종으로 분류하거나 다른 모든 곡비원류의 자매군의 일종으로 분류하는 이론이 제안되었다. 2008년에, 아이아이과(Daubentoniidae)는 여우원숭이하목이 되어야 하고, 뗏목을 타고 섬으로 건너 온 같은 선조 여우원숭이 개체군의 자손임이 확인되었으며, 아이아이과는 결과적으로 자신의 이름을 딴 아이아이하목에서 나와서, 여우원숭이하목에 속하게 되었다.[18]
단순한 코 또는 "마른 코"를 갖고 있는 영장류인, 직비원아목은 2개의 자매 분류군으로 이루어져 있다.[1]안경원숭이과(자신의 이름을 딴 안경원숭이하목에 속하는 단계통군)에 속하는 원원류 안경원숭이는 약 5,800만년 전에 가장 원시적인 부류를 대표한다.[26][27]
원숭이하목은 약 4,000만년 전에 등장했으며,[21] 2개의 분류군을 포함하고 있다: 남아메리카에서 발달했고 신세계원숭이를 포함하고 있는 광비원소목과 아프리카에서 발달하였고 구세계원숭이와 사람 등의 유인원을 포함하고 있는 협비원소목.[1] 세 번째 분류군, 에오시미드는 아시아에서 발달했지만 수백만 년 전에 멸종되었다.[28]
여우원숭이들처럼, 신세계원숭이도 기원이 명확하지 않다. 연쇄 상의 서열에 기반을 둔 분자적 서열 연구에서는 광비원류와 협비원류가 서로 분화된 때의 범위가 3,300 ~ 7,000만년 전으로 넓은 변이를 보이는 것으로 추정되는 반면에 미토콘도리아 염기 서열에 기반을 둔 연구에서는 좀 더 일치하는 3,500만년 전임을 시사하고 있다.[29] 진원류는 아프리카에 단일한 기원을 두고 있고 일부가 이동했음을 자명하게 받아들이고 있으며, 표본도 잇따라 발견되었다.[20] 진원류 영장류는 에오세 동안에, 대서양의 융기된 능선과 해수면이 낮은 바다가 이들의 이동을 쉽게 했을 것이며, 섬들을 징검다리로 하여 대서양을 건너 남아메리카에 도착했을 것으로 추정된다.[20] 한번 더, 일종의 뗏목 사건을 통해 이와 같은 대륙간 이주를 설명할 수 있을 것이다. 대륙 이동설에 의하면 처음 만들어진 대서양은 절대로 오늘날과 같이 넓을 수 없으며,[20] 연구 결과는 1 kg의 작은 영장류가 식물로 된 뗏목에서 어떻게 13일을 생존했는지를 보여주고 있다.[30] 해류와 풍속을 감안할 때, 대륙간을 항해할 충분한 시간이 제공되었다.

신세계원숭이의 일종, 황제타마린
유인원과 원숭이들은 마이오세가 시작될 무렵에 아프리카에서 유럽과 아시아로 퍼졌다.[31] 곧 이어, 로리스와 안경원숭이도 같은 여정을 떠났다. 가장 오래된 인류 화석은 500-800만년 전으로 거슬러 올라가며, 북아프리카에서 발견되었다.[21] 구세계원숭이들은 약 180만년 전에 유럽에서 사라졌다.[32] 일부 분자학적 연구와 화석 연구는 일반적으로 현세 인류가 10-20만년 전 아프리카에 기원을 두고 있다고 본다.[33]
영장류는 다른 동물군에 비해 연구가 잘 되고 있음에도 불구하고, 최근에도 몇몇 새로운 종들이 발견되고 있다(실험실에서의 유전학적 연구에 의해 이전에 불명확했던 종들의 구분이 밝혀지고 있다.). 2001년에 콜린 그로브스가 쓴 《영장류 분류(Primate Taxonomy)》는 약 350종을 수록하고 있으며,[34] 점점 증가하여 《세계의 포유류 종, Mammal Species of the World》(MSW3)의 제3판에는 376종이 수록되어 있다.[1] 그러나, MSW3는 현재 최종 2003년판에 알려진 일부 종들을 누락하고 있으며, 기록된 종의 수는 더 증가하여 424종을 기록하고 있고, 아종을 포함하면 658종에 육박하고 있다. 베마라하양털여우원숭이 (Avahi cleesei, 영국 배우이자 여우원숭이광인 존 클리세이의 이름을 땄다.)와 황금궁전티티원숭이를 포함하여 여러 새로 발견된 종들이 MSW3 목록에 누락되어 있다.
현존하는 영장류 분류
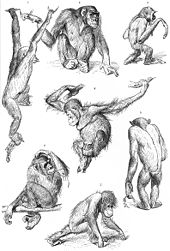
1927년의 침팬지, 긴팔원숭이(상단 오른쪽)와 오랑우탄 2마리(중앙과 하단 중앙) 그림. 상단 왼쪽은 나무가지를 건너뛰는 침팬지, 하단 중앙은 앞다리 지관절(指關節)의 등을 땅에 대고 걷는 오랑우탄
아래는 다양한 영장류의 목록이다.[1][19][35]
영장목 (PRIMATES) - 16과 495종
곡비원아목 (Strepsirrhini) - 7과 145종
여우원숭이하목 (Lemuriformes) - 5과 109종
난쟁이여우원숭이과 (Cheirogaleidae) - 40종
아이아이과 (Daubentoniidae) - 1종
여우원숭이과 (Lemuridae) - 22종
족제비여우원숭이과 (Lepilemuridae) - 27종
인드리과 (Indridae) - 19종
로리스하목 (Lorisiformes) - 2과 36종
로리스과 (Lorisidae) - 14종
갈라고과 (Galagidae) - 22종
직비원아목 (Haplorrhini) - 9과 350종
안경원숭이하목 (Tarsiiformes) - 1과 13종
안경원숭이과 (Tarsiidae) - 13종
원숭이하목 (Simiiformes) - 8과 337종
광비원류 (Platyrrhini): 신세계원숭이 - 5과 160종
비단원숭이과 (Callitrichidae) - 49종
꼬리감는원숭이과 (Cebidae) - 14종
올빼미원숭이과 (Aotidae) - 11종
사키원숭이과 (Pitheciidae) - 57종
거미원숭이과 (Atelidae) - 29종
협비원류 (Catarrhini) - 3과 177종
긴꼬리원숭이상과 (Cercopithecoidea): 구세계원숭이
긴꼬리원숭이과 (Cercopithecidae) - 151종
사람상과 (Hominoidea): 유인원 - 2과 26종
긴팔원숭이과 (Hylobatidae) - 18종
사람과 (Hominidae) - 8종

원원류이면서 직비원류로 간주되는 필리핀안경원숭이
영장목은 1758년에 린네에 의해, 그의 책 《자연의 체계》제10판[36] 에서 사람속(Homo, 사람), 시미아(Simia, 기타 유인원과 원숭이들), 여우원숭이속(Lemur, 원원류) 그리고 박쥐속(Vespertilio, 박쥐류)으로 명명되었다. 같은 책의 초판(1735년)에서, 린네는 사람속(Homo), 시미아속(Simia) 그리고 세발가락나무늘보속(Bradypus, 나무늘보)에 안드로포모르파(Anthropomorpha)라는 이름을 사용했다.[37]
1839년에 앙리 드 블랭빌은 린네의 명명법을 차용하여, Secundates목(Chiroptera, Insectivora, Carnivora아목을 포함), Tertiates목(또는 Glires), Quaternates목(Gravigrada, Pachydermata, Ruminantia을 포함)을 명명하였으나 [38] 이 새로운 분류는 채택되지 않았다.
1984년 앤더슨과 존스가 곡비원류와 직비원류라는 분류를 도입하기 전에(맥케나(McKenna)와 벨(Bell)의 1997년 저서 "포유류의 분류: 종 레벨 이상"에 의하면)[39], 영장류는 2개의 상과(원원류(Prosimii)와 진원류(Anthropoidea)로 나뉘었다.[40] 원원상과(Prosimii)는 모든 원원류를 포함(곡비원류에 안경원숭이를 더하여)했다. 진원상과(Anthropoidea)는 모든 진원류를 포함했다.
잡종
영장류 잡종은 사육 상태에서 종종 생기지만,[41] 야생에서 생기기도 한다.[42][43] 잡종은 2개 종의 활동 범위가 잡종 지역을 형성하여 겹칠 때 생긴다.; 잡종은 동물들이 동물원 등에 있을 때 인간에 의해 만들어지거나 약탈과 같은 환경적 압박때문에 만들어질 수 있다.[42] 속간 교잡 즉, 다른 속 사이의 잡종 또한 야생에서 발견되기도 한다. 비록 그들이 몇 백만 동안 구별되는 속에 속해 있지만, 이종 교잡은 단지 겔라다속과 망토개코원숭이 사이에서만 발생한다.[44]
구별되는 특징
영장류는 수상(樹上) 생활 서식지(나무와 관목)에 따라 다양한 종이 있으며, 이 환경에 적응하면서 생긴 다양한 특징을 갖고 있다.[45] 다음과 같은 구별되는 특징들이 있다.:
- 흉곽 안에 쇄골을 갖고 있다.[45]
- 모든 방향으로 큰 각도를 움직일 수 있는 어깨 관절이 있다[45]
- 엄지손가락과 나머지 손가락이 서로 마주 보게 할 수 있으며, 앞뒤 좌우 손과 발에 각각 5개씩의 손발가락을 갖고 있다.[45]
- 대부분의 종이 손톱과 발톱이 있다.[46]
- 현존하는 모든 종들은 엄지 손발가락의 손발톱이 평평하다.[46]
- 손가락 끝단에 민감한 촉각을 지닌 피부가 있다.[45]
- 눈구멍이 뼈로 둘러싸여 있다.[47]
- 주둥이가 짧고 얼굴이 평평한 편이며, 시각에 비해 후각은 떨어진다.(대부분의 직비원류가 그렇고, 곡비원류는 덜한 편이다).[47]
- 입체상이 가능하고 높은 시각과 색각을 갖춘 복잡한 시각 인식 체계를 갖고 있다.[45]
- 뇌는 잘 발달된 후엽소뇌와 새발톱고랑(조거구)를 갖고 있다.[47]
- 몸 크기에 비해 상대적으로 큰 뇌를 갖고 있다. 유인원은 특히 큰 뇌를 갖고 있다.[45]
- 대뇌 피질이 더 크게 분화되어 있다.[45]
- 원시 포유류보다 치아 개수가 상대적으로 적다.[45]
- 치아가 3종류로 나뉜다.[47]
맹장이 잘 발달되어 있다.[47]- 2개의 유선(乳腺)이 있다.[45]
- 보통 한 번에 한 마리의 새끼를 밴다.[45]
- 수컷 생식기와 고환이 흔들거린다.[47]
- 임신 기간과 발생 기간이 길다.[45] 그리고
- 몸통을 똑바로 유지하려고 하며, 이것이 두 발 보행을 하게 만든다.[45]
모든 영장류가 이런 해부학적인 특징을 보여주는 것도 아니며, 어느 특징도 영장류에만 있는 것도 아니다. 예를 들어, 다른 포유류들도 쇄골과 3종류의 치아와 흔들거리는 수컷 생식기를 가지고 있는 반면에, 거미원숭이들은 매우 줄어든 엄지손가락을 지니고 있으며, 목도리여우원숭이들은 6개의 유선이 있고, 원원류는 일반적으로 코가 길고 냄새를 아주 잘 맡는다. 영장류는 종합적인 포유류다.[47]
행동 면에서, 영장류는 높은 수준의 사회적 행동을 종종 하며, 유연하고 지배적인 위계 체계를 갖고 있다.[48] 신세계원숭이는 대부분의 구세계원숭이와는 달리, 단혼의 일자일웅 형태를 취하며, 수컷이 새끼를 돌본다.[49]
해부학, 생리학 그리고 형태학
영장류는 두개골 전면의 얼굴에 앞을 향한 눈을 지니고 있다; 양안시는 거리를 정확하게 지각하고, 모든 유인원의 선조들이 나무가지를 팔로 매달리며 건너다니기 유용하도록 해주었다.[45] 안와 위에 일종의 뼈가 융기된 곳이 있다; 이 융기된 곳은 씹는 동안에 피로가 쌓이는 얼굴 내의 약한 뼈들을 보강해 준다. 곡비원류는 눈을 보호하기 위해 안와 둘레에 일종의 후안와봉을 가지고 있다; 반면에, 더 고등한 영장류인, 직비원류는 완전히 진화된 안와를 지니고 있다.[50]

영장류 두개골 크기 및 무게 비교
영장류 두개골은 둥글고 큰 덮개를 가지고 있으며, 특히 안드로포이드(anthropoid)에게 두드러진 특징이다. 두개골 덮개는 이 집단의 구별되는 특징의 하나인, 큰 뇌를 보호하는 역할을 한다.[45] 뇌용적(두개골 내의 부피)은 일종의 뇌 크기를 반영하며, 사람이 나머지 영장류들보다 세 배나 크다.[51] 평균 뇌용적은 사람이 1201 cm3, 고릴라가 469 cm3, 침팬지가 400 cm3 그리고 오랑우탄이 97 cm3이다.[51] 영장류 진화의 주요 방향은 두뇌의 정교화에 있으며, 특히 감각 인지와 운동 명령의 생성, 공간 추론, 의식적인 사유 그리고 사람의 언어와 관련이 있는 신피질 (대뇌 피질의 일부분)에 있다.[3] 다른 포유류들이 후각에 상당히 의존하는 반면에, 영장류들의 수상(樹上) 생활은 일종의 촉각과, 시각을 지배적인 감각 기관으로 이끌었으며,[3] 뇌에서 후각을 담당하는 부분의 축소와 복잡한 사회적 행동의 증가를 낳았다.[52]

영장류의 손과 발을 그린 1893년의 그림
영장류는 원래 각 손발에 5개의 손발가락(五指)을 가지고 있으며, 각 손발가락의 끝에는 각질로 된 손발톱이 있다. 손발의 바닥 면은 손발의 끝에 민감한 피부를 지니고 있다. 대부분 영장류의 특징 중 하나인 마주보게 할 수 있는 손가락을 가지고 있다. 그러나 마주보게 할 수 있는 손가락은 영장류만 있는 것은 아니다(예를 들어, 주머니쥐도 가지고 있다.).[45] 일부 종들은 엄지손가락을 통해 도구를 사용할 수 있다. 영장류에서, 마주보게 할 수 있는 손가락들의 배합, 즉 짧은 손발톱(고양이 매 따위의 발톱이기 보다는)과 길고 안쪽으로 접히는 손가락들은 나무 가지를 움켜쥐던 습성의 영장류 선조들의 유물 중 하나이며, 부분적으로, 일부 종들에게 매우 중요한 이동수단으로써 팔로 매달리며 건너다닐 수 있는 기술을 발달시켜 주었다. 원원류는 각 발의 두 번째 발가락에 고양이 매 따위의 발톱과 같이 생긴 발톱이 있으며, "몸단장용-발톱"이라고 부르며, 털을 손질할 때 사용한다.[45]
영장류의 쇄골은 팔이음뼈의 두드러진 요소로써 유지되어 왔으며, 이는 어깨 관절의 운동성을 폭넓게 만든다.[48] 유인원은 등쪽에 위치한 어깨뼈와 앞뒤로 평평하고 넓은 흉각 때문에 어깨 관절과 팔을 더 잘 움직일 수 있으며, 구세계원숭이와 비교하여(아랫쪽 척추뼈가 크게 감소하고, 일부 종에서 꼬리가 사라진) 더 짧고, 운동성이 떨어지는 등뼈를 가지고 있다. 구세계원숭이는 유인원과는 달리 대부분 꼬리를 지니고 있다. 뭔가를 쥘 수 있는 꼬리를 지닌 영장류는 신세계원숭이인 고함원숭이와 거미원숭이 그리고 양털원숭이 등을 포함하고 있는 거미원숭이과 원숭이가 유일하다.
영장류는 진화적으로 주둥이가 줄어드는 경향을 보여주고 있다.[48] 기술적으로, 구세계원숭이는 코의 구조에서 신세계원숭이와 구별되며, 치열에서 유인원과 구별된다.[52] 신세계원수이의 콧구멍은 얼굴 옆으로 향해 있으며, 구세계원숭이는 얼굴 아래쪽으로 향해 있다.[52] 영장류의 치아 형태는 상당히 다양하며, 일부는 앞니가 거의 없는 반면에, 모두 최소한 아래 앞니 하나는 유지하고 있다.[52] 대부분의 곡비원류에서, 아래 앞니와 송곳니로 일종의 참빗을 형성하여, 털을 손질하거나 때로는 먹이를 구하는 데 사용하며,[47][52] 첫 번째 아래 어금니는 송곳니와 같은 형상을 하고 있다.[47] 구세계원숭이는 8개의 작은어금니를 지니고 있어, 12개를 지닌 신세계원숭이와 비교된다.[52] 유인원은 5개, 구세계원숭이는 4개이며,[52] 반면에 사람은 4개 또는 5개이다.[53] 중요한 호미니드 어금니 교두(hypocone)는 영장류 역사 초기에 진화한 반면에, 이에 상응하는 아래 어금니(파라코니드)의 교두는 퇴화했다. 원원류는 움직이지 않는 고정된 윗 입술을 갖는 특징이 있으며, 코 끝이 젖어 있고 아래 앞니가 앞으로 나와 있다.
영장류에 있어서 색각의 진화는 다수의 진수하강 포유류 중에서 독특하다. 영장류의 먼 척추동물 조상이 3색성 색각을 보유한 반면에, 야행성이자 온혈 동물인 포유류 조상은 중생대 기간 동안에 망막에서 3개의 추체 중의 하나를 상실했다. 어류와 파충류 그리고 조류는 그러므로 3색성 색각 또는 4색성 색각을 가지고 있는 반면에, 일부 영장류와 유대류[54] 를 제외한 모든 포유류는 2색성 색각 또는 단색성 색각 (완전히 색을 분별하지 못하는)을 가지고 있다.[47]올빼미원숭이와 부시베이비와 같은 야행성 영장류는 흔히 단색성 색각을 가지고 있다.
협비원류는 3~4천 만년 전, 그들의 조상 단계에서 적록 옵신 유전자가 유전자 복제되었기 때문에 3색성 색각일 수밖에 없다.[47][55] 한편, 광비원류는 몇몇 경우만이 3색성 색각이다.[56] 엄밀히 말하면, 개개의 암컷들은 X 염색체의 같은 자리에 위치하는 옵신 유전자(적록)의 2개의 대립 유전자가 이형이어야 한다.[47] 수컷은 그러므로 단지 2색성 색각일 뿐이지만, 반면에 암컷은 2색성 색각이거나 3색성 색각일 수 있다. 또한 곡비원류는 색채를 식별할 수 없다. 그러나 연구에 의해, 광비원류에게서 발견되는 것과 비슷한, 일정한 범위의 색각이 있음이 지적되고 있다.[47]
협비원류처럼, 고함원숭이(광비원류의 일종)는 진화 상 최근에 유전자 복제된 것으로 밝혀진 전형적인 3색성 색각을 보여 준다.[57] 고함원숭이는 신세계원숭이 중에서 잎을 먹는 행동이 가장 발달한 원숭이 중 하나이다. 과일이 이들 먹이의 주 먹이가 아니며,[58] 이들이 먹는 잎(어리고, 영양이 있고, 소화가 잘 되는)의 종류는 단지 일종의 적록 신호에서만 찾아 낼 수 있다. 고함원숭이가 선호하는 먹이를 탐색하는 현지 조사는 전형적인 3색성 색각이 환경적으로 선택된 것임을 암시하고 있다.[59]
성적 이형성

크기가 뚜렷하게 차이가 나는 망토개코원숭이 수컷(회색)과 암컷(갈색) 사이의 성적 이형성
같은 종에서 다른 성의 개체 사이의 차이인, 성적 이형성(동종이형)이 진원류에서 흔히 나타나며, 신세계 종들보다는 구세계 종들(유인원과 일부 원숭이) 사이에서 그 차이가 더 큰 것으로 보인다. 최근의 연구는 영장류에서 나타나는 이형성의 변이와 성적 이형성의 근본 원인, 둘 다를 조사하기 위하여 DNA 비교 방법을 쓴다. 영장류는 흔히 몸무게[60][61] 와 송곳니의 크기[62][63], 그리고 털와 피부의 색깔에서 성적 이형성을 가지고 있다.[64] 이와 같은 성적 이형성은 짝짓기 방식,[65] 크기,[65] 서식 환경과 먹이를 포함한 다양한 요인에 의해 영향을 받거나 영향을 끼친다.[66]
비교 분석을 통해 영장류의 성 결정, 자연 선택 그리고 짝짓기 방식 사이에서의 관계에 대하여 좀 더 완벽하게 이해할 수 있게 되었다. 연구는 성적 이형성이 암수의 특징의 변화를 만들고 있음을 보여 주고 있다.[67] 일반적인 성장 과정에서의 비례적인 신장이 일어나는, 발생적 크기 조정은 성적 이형성과 성장 형태 사이의 관계에 대해 약간의 통찰을 제공하는 것으로 보인다.[68] 화석 기록에서의 일부 증거는 성적 이형성의 수렴 진화를 암시하고 있으며, 일부 멸종된 인류는 현존하는 영장류보다 성적 이형성이 더 큰 것으로 보인다.[67]
이동

다른 나무로 건너 뛰려하는, 수직으로 매달리고 건너뛸 수 있는 왕관시파카
영장류 종은 팔로 매달리며 건너다니기, 두발보행, 뛰어오르기, 나무 위와 지상 에서의 네발보행, 기어오르기, 앞다리의 지관절(指關節)의 등을 땅에 대고 걷기를 통해 이동하거나 이 방법들을 조합하여 이동한다. 몇몇 원원류는 주로 수직으로 매달리거나 뛰어 오르기를 한다. 이들에는 부시베이비의 다수와 모든 인드리 (예들 들어, 시파카, 아바히 그리고 인드리), 족제비여우원숭이, 그리고 모든 안경원숭이를 포함된다.[69] 나머지 진원류는 나무 위에서 네발을 사용해 움직이거나 나무를 타고 오른다. 일부는 또한 지상에서 네발로 걷거나 일부는 뛰어 다닌다. 대부분의 원숭이는 나무 위에서 생활하거나 지상에서 네발로 걷거나 기어오르기 둘 다를 한다. 긴팔원숭이, 무리키 그리고 거미원숭이들 모두 "팔로 매달려 건너다니기"를 광범위하게 사용한다.[32]양털원숭이들 또한 종종 두팔로 매달려 건너다닌다.[58]오랑우탄은 일종의 "사지형 기어오르기"라고 불리는 비슷한 방식으로 이동하는 데, 이 방식은 나무들 속에서 자신의 무거운 몸을 이동시킬 때, 자신의 팔과 다리를 사용하는 것이다.[32]침팬지와 고릴라는 앞다리의 지관절(指關節)의 등을 땅에 대고 걸으며,[32] 짧은 거리는 두발로 걸을 수 있다. 오스트랄로피테신과 초기 인류와 같은 수많은 종들이 완전히 두발로 이동했음이 밝혀져 있지만, 사람만이 이러한 특징을 갖는 현존하는 유일한 종이다.
습성
사회 체계
리처드 랭햄은 사람을 제외한 영장류의 사회 조직은 집단 내에서 암컷이 움직이는 양에 따라서 가장 잘 분류할 수 있다고 주장했다.[70] 그가 제안한 4가지 범주는 다음과 같다.:
- 암컷 전출 조직 – 암컷은 자신이 태어난 집단을 떠난다. 한 집단 내의 암컷들은 친밀한 관계가 없는 반면에 수컷들은 자신이 태어난 집단에 남는 데, 이런 밀접한 연관성은 사회적 행동에 영향을 받는 것으로 보인다. 이런 집단은 일반적으로 매우 작은 집단을 형성한다. 이런 조직은 침팬지에서 발견되며, 대표적으로 관련이 있는 수컷들은, 자신들 영역을 지키기 위해 협력할 것이다. 신세계원숭이들 중에서는, 거미원숭이들과 무리키들이 이런 체계를 사용한다.[71]

지고쿠다니 온천에서 함께 목욕하는 일본마카크들
- 수컷 전출 조직 – 암컷들이 자신이 태어난 집단에 머무는 반면에, 수컷들은 어렸을 때 집단을 떠난다. 일부다처제와 다부 사회들이 이 범주에 속한다. 통상적으로 집단의 크기가 크다. 이 조직은 호랑이꼬리여우원숭이, 카푸친원숭이들 그리고 긴꼬리원숭이아과 원숭이들 중에서 흔하다.[32]
- 일부일처제의 종 – 한 마리의 수컷-암컷의 결합, 때로는 청년기의 새끼를 동반하기도 한다. 새끼들 돌보기와 영역을 지키는 책임을 함께 진다. 새끼는 청년기 동안에 부모의 영역을 떠난다. 긴팔원숭이는 본래 이 체계를 사용하며, 그렇지만 이 맥락에서 "일부일처제"는 반드시 절대적인 성적인 적합성을 의미하지는 않는다.[72]
- 군거 생활을 하지 않는 종 – 흔히 수컷들이 몇몇 암컷들의 서식 범위를 포함하는 영역을 지킨다. 이 종류의 조직은 진원류에서 발견된다. 오랑우탄은 자신의 영역을 고집하지는 않지만, 실제로는 이 조직을 가지고 있다.[73]
다른 조직들도 발생하는 것으로 알려져 있다. 예를 들어, 고함원숭이들을 성적으로 성숙기에 도달하면 암수 모두 자신이 태어난 집단에서 떠나기 때문에, 결과적으로 집단 내에서 수컷이든 암컷이든 전형적인 관계는 없다. 탄자니아 곰비 천 국립공원에서 연구 활동을 한 영장류 동물학자 제인 구달은 침팬지들 내의 분열-융합 사회 구조를 주목했다.[74] 주요 집단은 낮 동안에 먹이를 찾기 위해 분리되는 "분열"을 한 다음에, 밤에는 잠을 자기 위해 한 집단 내로 돌아 오는 "융합"을 한다. 이런 사회 구조는 망토개코원숭이[75] 와 거미원숭이[58], 보노보[75] 에서도 발견될 수 있다. 겔라다는 비숫한 사회 구조를 가지는 데, 많은 작은 집단들이 함께 600마리까지 임시적인 집단을 형성한다.[75]
이들 사회적 체계는 다음 3개의 주요 생태 환경 요인에 의해 영향을 받는다.: 자원의 분포, 군집의 크기 그리고 포식.[76] 특정한 군집 내에는 협조와 경쟁사이에 일정한 균형이 있다. 협조 행동은 사회적 그루밍(피부의 기생충 제거 및 상처 손질), 먹이 공유하기, 그리고 맹수로부터 일정한 영역 내 또는 영역에 대한 집단적인 방어를 포함한다. 적대 행동은 가끔 이용할 수 있는 먹이나 잠자는 곳 또는 짝에 대한 경쟁을 표시하기도 한다. 공격 행동은 또한 지배적 지위를 얻기 위한 용도로 사용된다.[49][77]
이종간 군집
영장류의 몇몇 종들은 야생에서 서로 연합하는 것으로 알려져 있다. 이들 연합의 일부는 광범위하게 연구되어 왔다. 아프리카의 타이 숲에서, 몇몇 종들은 포식동물로부터 방어하기 위해 협력하는 습성이 있다. 이들 종은 다이애나원숭이, 캠벨모나원숭이, 작은흰코원숭이, 서부붉은콜로부스, 왕콜로부스 그리고 검댕망가베이이며, 포식 동물로부터 방어하기 위한 경고 소리를 통해 협력한다.[78] 이 원숭이들을 노리는 포식동물은 침팬지이다.[79]
붉은꼬리원숭이는 서부붉은콜로부스, 푸른원숭이, 울프모나원숭이, 망토게라자, 검은볏망가베이, 앨런습지원숭이를 포함한 여러 종들과 연합한다.[75] 이들 종 몇몇은 침팬지에 의해 잡아 먹힌다.[80]
남아메리카에서, 다람쥐원숭이들은 카푸친원숭이들과 연합한다.[81] 다람쥐원숭들에게 포식동물을 방어하는 것보다 먹이를 찾는 일에 더 도움이 되는 것으로 보인다.[81]
인지 능력과 의사 소통
영장류는 진일보한 인지 능력을 지니고 있다. 일부는 도구를 만들어, 먹이를 구하고 사회적 표현을 하는 데 사용한다.[82][83] 일부는 협력과 힘 그리고 지위 등이 필요한 지능적인 사냥 전략을 구사한다.[84] 이들은 자신의 지위를 인지하고, 꾸미거나 속일 수 있는 능력이 있다.[85] 자신들과 같은 종인지 다른 종인지도 인식할 수 있으며,[86][87] 기호를 사용하는 법을 배우고, 일부의 문법 규칙과 수와 순서의 개념을 포함한 인간 언어의 양상을 이해할 수 있다.[88][89][90] 영장류 인지에 대한 연구는 문제 해결, 기억, 사회적 상호작용, 일종의 마음 이론, 그리고 수리 및 공간 지각 그리고 추상적 개념 등을 탐구한다.[91]
여우원숭이, 로리스, 안경원숭이 그리고 신세계원숭이들은 사회적, 생식 행동의 여러 국면에서 후각 신호에 의존한다.[3] 분화된 분비선인 페로몬을 자신의 영역을 표시하기 위해 사용하며, 서골비기관을 통해 탐지한다. 이 과정이 이들 영장류의 의사 교환 행동 중에서 가장 큰 부분이다.[3] 구세계원숭이들과 유인원들에 대부분 흔적으로만 남아 있고 삼색형 색각의 시력으로써 퇴행한 이 능력이 주 감각 기관이 되도록 진화했다.[92] 영장류는 또한 심리 상태를 전달하기 위해 발성, 몸짓, 얼굴 표현을 사용한다.[93]
생활사
영장류는 다른 포유류들보다 발달이 늦다.[32] 사람을 제외한 모든 영장류의 새끼들은 어미가 모유 수유를 하며, 돌보고 데리고 다닌다.[32] 일부 종들에서는, 그 집단 내의 수컷들이 새끼들을 보호하며, 데리고 다니며, 특히 그 수컷들은 그들의 아버지로 보인다.[32] 형제 자매와 아주머니와 같은 새끼들의 기타 친척들 또한 이들을 보살필 것이다.[32] 대부분의 영장류 어미들은 새끼들의 모유 수유 기간 동안에는 배란이 중단된다. 그 새끼들이 젖을 떼면 그 어미는 다시 생식을 할 수 있다.[32] 이는 젖을 계속 먹으려는 새끼와 젖떼기 갈등을 일으키는 원인이 되기도 한다.[32]
영장류는 비슷한 크기의 다른 포유류에 비해, 젖을 떼고 성적으로 성숙해지는 기간이 길다.[32] 성적으로 성숙해지기 이전 기간 동안에, 영장류는 포식과 굶주림에 더 민감하다. 이 시기 동안에 이들은 먹이를 구하고 맹수를 피하는 방법을 경험을 통해 얻는다.[32] 또한 이들은 종종 놀이를 통해 군집 생활 및 싸움 기술을 배운다.[32]
영장류, 특히 암컷들은, 비슷한 크기의 다른 포유류보다 수명이 더 길다.[32]
먹이

흑백콜로부스의 일종으로 잎을 먹고 있는 망토게레자

먹이를 구해 자신의 볼주머니에 임시로 저장하는 게잡이마카크
영장류는 여러 가지의 음식의 공급원을 개발한다. 사람을 포함하여, 현대 영장류의 많은 특징들은 열대 우림의 캐노피에서 이들 먹이의 대부분을 구하던 초기 선조들의 습성에서 유래한 것이라고 말해지고 있다.[94] 대부분의 영장류들은 에너지를 얻기 위해, 탄수화물과 지질이 쉽게 소화되는 과일을 이들의 먹이에 포함시킨다.[32] 그러나 아미노산, 비타민 그리고 미네랄을 얻기 위해, 잎 또는 곤충과 같은 기타 음식들도 필요로 한다. 많은 영장류들은 과일, 잎, 수지(樹脂) 또는 곤충 등과 같은 특이한 음식들을 섭취하기 위하여 해부학적으로 분화된 기관들을 지니고 있다.[32] 예를 들어, 고함원숭이, 흑백콜로부스와 족제비여우원숭이처럼, 잎을 먹는 원숭이들은 소화시키기 어려울 수 있는 잎에 있는 영양분을 흡수하기 위하여 확장된 소화관을 지니고 있다.[32] 수지를 먹는 동물인 마모셋은 수지를 꺼내기 위해 나무껍질을 벗길 수 있도록 강한 앞니가 있으며, 음식을 먹는 동안 나무에 매달릴 수 있도록 사람의 손발톱보다는 맹금류와 같은 발톱을 지니고 있다.[32]아이아이는 설치류처럼 길고 가는 가운뎃손가락을 지니고 있으며, 이는 딱다구리의 생태학적 지위와 같은 역할을 한다. 곤충의 유충을 찾기 위해 나무들을 두드리고, 그 다음에 나무를 갉아 구멍을 내고 유충을 꺼내기 위해 가운뎃손가락을 구멍에 집어넣는다.[95] 일부 종들은 부가적인 분화 기관을 가지고 있다. 예를 들어, 회색뺨망가베이는 이빨에 얇은 법랑질이 있어, 다른 원숭이들이 열 수 없는 딱딱한 과일과 종자들을 열 수 있다.[32]
겔라다는 풀을 주식으로 하는 유일한 영장류다.[96]안경원숭이는 육식을 하는 유일한 영장류로, 오로지 곤충과 갑각류, 작은 척추동물과 뱀들(독사 포함)만을 먹는다.[97] 이와 반대로 카푸친원숭이들은 과일, 잎, 꽃, 새싹, 과즙, 씨앗, 곤충 그리고 기타 무척추동물과 새알, 그리고 새들과 도마뱀, 다람쥐, 박쥐와 같은 작은 척추동물들을 포함하여 여러 종류의 음식들을 섭취할 수 있다.[58]침팬지는 서부붉은콜로부스 원숭이와 같은 다른 영장류를 잡아 먹는 것을 포함하여 다양한 음식을 먹는다.[98][99]
서식지 및 분포

인도 아그라 성의 붉은털원숭이
영장류는 수상(樹上) 동물로부터 진화했으며, 많은 종들이 대부분의 시간을 나무 위에서 생활하고 있다. 대부분 열대 우림에서 산다. 열대기후 지역에 사는 영장류 종의 수는 강수량 그리고 우림의 면적과 양의 상관 관계가 있는 것으로 보인다.[100] 열대 우림에서 과일을 상식하는 동물들(무게 기준) 중에서 25~40%를 차지하는 영장류는 여러 수종의 씨앗을 퍼뜨리는 데 생태학적으로 중요한 역할을 한다.[101]
개코원숭이와 파타스원숭이와 같은 일부 종들은 부분적인 영역과시형 동물이며, 겔라다와 사람과 같은 소수의 종들은 완전한 영역과시형 동물들이다. 사람을 제외한 영장류는 우림과 망그로브 숲 그리고 저산대 숲을 포함하여 아프리카, 인도, 동남아시아, 그리고 남아메리카의 열대 기후 지역 위도 상의 여러 숲 서식지에서 산다. 비-영장류의 일부는 열대 지방이 아닌 것에서 서식한다. 예를 들어, 산악 지역에 사는 일본원숭이는 연중 8개월 이상이 눈으로 덮여 있는 혼슈 섬 북쪽에 살며, 바르바리마카크는 알제리와 모로코의 아틀라스 산맥에서 산다. 영장류는 일정한 범위의 고도에서 서식한다.: 검은들창코원숭이는 해발 4,500미터의 헹두안 산맥에서 사는 것이 발견되고 있으며,[102]산악고릴라는 비룽가 산맥을 횡단하는 해발 4,200미터 고도에서도 발견될 수 있으며,[103] 겔라다원숭이는 에티오피아 고지의 해발 5,000미터 높이에서 발견되고 있다.
대부분의 종들이 일반적으로 물을 두려워하지만, 코주부원숭이, 브라자원숭이 그리고 알렌습지원숭이를 포함하여 일부 종들은 훌륭한 수영선수이며 습지와 물이 있는 지역에서 편안함을 느끼고 손가락 사이에 작은 물갈퀴 막이 발달해 있다. 붉은털원숭이와 회색랑구르와 같은 일부 영장류들은 인공적인 환경을 이용할 수도 있으며, 심지어 도시에서도 산다.[75][104]
대부분이 온대 기후까지밖에 살지 않는데 비해서 사람은 추운 지방에서도 살고 있다.
사람과의 상호 작용
일부는 사람과 사람 이외의 영장류 사이의 밀접한 관계와 상호 작용이 동물원성(原性) 감염 질병이 전염되는 경로를 만들어 내고 있다는 가설을 세우고 있다. 헤르페스 바이러스(가장 흔하게는 헤르페스 B 바이러스)와 같은 바이러스, 천연두 바이러스, 홍역, 에볼라, 광견병, 마르부크 바이러스 그리고 바이러스성 감염 등은 사람들을 감염시킬 수 있다. 일부 경우에, 바이러스는 사람 및 사람을 제외한 영장류 모두에게 치명적인 질병을 일으킬 가능성이 있다.[105]
법적 사회적 상태
오직 인간만이 개인으로 인정되고, 국제 연합 세계 인권 선언에 의한 법으로 보호되고 있다. 한편, 사람 이외 영장류에 대한 법적 지위는 적어도 그들 중 일부에게는 법적 권리를 부여해야 한다고 운동을 벌이는 그레이트 에이프 프로젝트(Great Ape Project, GAP)와 같은 단체들과 논쟁을 많이 벌이는 주제이다.[106] 2008년 6월, 스페인은 그레이트 에이프 프로젝트(GAP)의 권고에 호응하여, 의회의 여러 정당이 참가한 환경위원회가 정부에 촉구함으로써, 세계에서 사람이 아닌 영장류의 권리를 인정한 첫 번째 국가가 되었으며, 그 내용은 침팬지와 보노보, 오랑우탄 그리고 고릴라는 동물 실험의 대상이 될 수 없다라는 것이다.[107][108]

카푸친원숭이는 손발을 능숙하게 쓸 수 있기 때문에 사지가 마비된 사람들을 도울 수 있다.
많은 수의 사람이 아닌 영장류들이 사람들에 의해서 애완용으로 길러지고 있다. 그레이트 에이프 프로젝트는 약 3,000마리의 사람이 아닌 영장류들이 미국에서 애완용으로 살고 있다고 추산하고 있는 반면에, 미국 휴메인 소사이어티는 더 높게 잡아서 약 15,000마리로 보고 있다.[109] 최근 수년동안, 중국의 중산층이 성장함에 따라서 색다른 애완동물로써 사람이 아닌 영장류에 대한 수요가 증가하고 있다.[110] 1975년 미국이 영장류를 애완용으로 수입하는 것을 금지하였음에도 불구하고, 여전히 미국-멕시코 국경을 통해 영장류가 밀수입되고 있으며, 원숭이는 미화로 3000달러에 유인원은 30,000달러에 거래되고 있다.[111]
영장류는 실험실에서 모델 생물로 사용되며, 우주 비행을 한 적도 있다.[112] 불구가 된 사람들을 위한 서비스 동물로도 일한다. 꼬리감는원숭이는 사지가 마비된 사람들을 돕도록 훈련되기도 한다. 이들의 지능과 기억력 그리고 손발을 쓰는 능력은 훌륭한 도우미가 되게 한다.[113]
사람이 아닌 영장류들은 전 세계 동물원에서 사육되고 있다. 역사적으로, 동물원들은 주 기능이 오락의 일종이었지만, 좀 더 최근에는 보존과 교육, 연구로 그 초점이 옮겨졌다. 많은 동물원들은 이제, 공공에 자연 전시와 교육 자료로써 주로 이용된다. 미국에서 많은 단체들이, 사육 상태의 번식을 통한 유전적 다양성을 극대화시키기 위해 미국 동물원 수족관 협회(AZA)에서 개발한, 종 생존 계획(SSP)에 참가하고 있다. 동물원들과 기타 동물 복지 지지자들은 일반적으로 동물 권리 우선권과 모든 사람이 아닌 영장류들이 사육 상태에서 벗어나야 한다는 GAP의 주장을 크게 두 가지 이유로 반대한다. 첫째, 사육 상태에서 태어난 영장류들은 풀려났을 때, 야생에서 생존할 수 있는 지식과 경험이 없다. 둘째, 동물원은 야생에서 멸종 위험에 처한 영장류와 기타 동물들에게 생활 공간을 제공하고 있다.
과학 연구에서의 역할
수천 마리의 사람이 아닌 영장류들이 인간과 심리적 생리적으로 유사하기 때문에 전 세계에서 과학 연구에 사용되고 있다.[114][115] 특히, 사람이 아닌 영장류의 뇌와 눈은 어느 다른 동물들의 그것보다 사람과 해부학적으로 좀 더 유사하다. 사람이 아닌 영장류들은 신경과학, 안과 연구, 그리고 독성 연구를 위한 임상시험 용으로 흔히 사용된다. 히말라야원숭이도 자주 사용되며, 야생에서 포획하여 목적을 가지고 기르는 마카크와 아프리카 푸른원숭이, 침팬지, 개코원숭이, 다람쥐원숭이, 그리고 마모셋 등도 사용된다.[114][116] 2005년에, GAP는 미국에서 사는 3100마리의 사람이 아닌 영장류의 1280마리가 실험용으로 사용되었다고 보고했다.[106] 2004년, 유럽 연합은 그와 같은 실험에 약 10,000마리의 사람이 아닌 영장류가 사용했다. 2005년 영국은 실험 동물 4,652 마리 중 3,115 마리의 사람 아닌 영장류가 실험용으로 사용되었다.[117] 많은 나라의 정부는 사육 상태에 있는 사람이 아닌 영장류에 대한 엄격한 관리 요구 조건을 가지고 있다. 미국에서, 연방 정부 지침으로 사람이 아닌 영장류의 주거, 식생활, 영양 그리고 번식에 대한 견해를 포괄적으로 규정하고 있다.[118] 동물 실험을 중지시키기 위한 '유럽 연합'과 같은 유럽의 단체들은 유럽연합의 동물실험 입법의 재검토의 일환으로써 실험에 사용되는 모든 '사람이 아닌 영장류'의 실험을 금지시키는 법안을 모색하고 있다.[119]
보존
국제 자연 보호 연맹(IUCN) 목록은 영장류의 1/3 이상을 멸종위급 또는 취약으로 분류하고 있다. 영장류 종들에 대한 공통적인 위협은 무분별한 벌목과 산림 파편화, 원숭이 쫒기(영장류의 곡식 피해로 인한)가 포함되며,[120] 의약품과 애완동물 그리고 음식으로 사용되기 위한 영장류 사냥 등이 있다. 대규모의 열대 지역 산림 개간이 영장류 멸종 위협의 가장 큰 요인으로 간주되고 있다.[121][122][123] 영장류 종의 90% 이상이 열대 지역 숲에 산다.[122][124]
숲 감소의 요인으로 상업 벌목, 생계형 목재 채취, 탄광 채굴 등이 있지만, 주 요인은 농업을 위한 개간이며, 댐 건설 또한 열대 숲의 수원지 고갈을 일으킨다.[124]팜유 생산을 증대시키기 위해, 인도네시아 저지대 우림 지역을 대규모로 개간하여 왔으며, 위성 사진을 분석한 결과, 1998년과 1999년 사이에 레우서르 밀림에서만 1년에 1,000여 마리의 수마트라오랑우탄이 사라졌다는 보고가 나왔다.[125]

멸종위급종인 수마트라오랑우탄
몸집이 큰 영장류(5 kg 이상)는 작은 영장류와 비교하여 밀렵꾼에게 붙잡힐 가능성이 높기 때문에 멸종 위험이 증가한다.[124] 영장류는 다른 동물들보다 성숙이 늦은 편이며, 새끼를 낳는 간격이 길다. 그러므로 밀렵이나 애완동물 거래 등으로 개체수가 한 번 감소하면, 회복되는 속도가 늦다.[126] 일부 아프리카 도시에서 보고되는 정보에 의하면, 도시 지역에서 소비되는 모든 단백질의 절반 정도가 부시미트 무역에서 생긴다.[127]게논과 드릴원숭이와 같은 멸종 위험종 영장류는 지속가능한 상태를 크게 초과하는 수준에서 사냥이 되고 있다.[127] 이것은 몸 크기가 크고, 운반이 쉬우며, 동물 개체 당 이익률이 높기 때문이다.[127] 농업이 숲 서식지까지 파고들고 있고, 영장류가 그 곡식들을 먹기 때문에, 농부들이 경제적으로 큰 손실을 보게 만들고 있다.[128] 영장류가 농작물을 망치는 행위는 현지에서 영장류에 대한 보존 노력을 방해하는 부정적인 인식을 심어주고 있다.[129]
가장 광범위한 멸종은 영장류 과 중에서 4개 과의 원주지인 마다가스카르에서 발생했다. 사람들이 살기 시작한 1,500여 년 전 이래, 서식지 파괴에 의해 적어도 8개 강과 15개 종이 멸종했다.[130] 멸종된 영장류 중에는 Archaeoindris속 (산악고릴라보다 큰 여우원숭이의 일종)과 Palaeopropithecidae과, Archaeolemuridae과가 있다.[3]
아시아에서, 힌두교와 불교 그리고 이슬람교는 영장류 고기를 먹는 것을 금지하고 있다. 그러나 영장류는 아직도 식용으로 사냥되고 있다.[124] 일부의 규모가 적은 전통 종교는 영장류 고기의 소비를 허용하고 있다.[131][132]애완동물 거래와 전통 의약품 또한 불법적인 사냥이 증가하는 원인이다.[110][133][134]모델 생물의 일종인 히말라야원숭이는 1960년대에 그 수를 위협하는 과도한 사냥 이후에 사냥을 금지하였다. 이 프로그램은 효과가 너무 커서, 이제는 이 마카크가 전 분포 지역에서 골칫거리가 되고 있다.[123]

멸종위급종 크로스강고릴라
중앙아메리카와 남아메리카에서, 숲 파편화와 사냥은 영장류에서 가장 중요한 2개의 문제이다. 광대한 넓이의 숲은 이제 중앙아메리카에서 드물게 되었다.[121][135] 이는 농장의 잠식, 낮은 레벨의 습도, 식물 식생의 변화처럼 주변 효과에 취약한 숲의 크기를 키운다.[136][137] 운동의 제한은 큰 폭의 근친교배의 증가를 초래하여, 개체군이 현저하게 사라지는 일종의 개체군 병목현상을 이끌어, 저하 효과를 일으킬 수 있다.[138][139]
현재 멸종위급종 영장류는 21종이며, 2000년에 발표된 IUCN의 ‘세계에서 멸종 위험이 가장 높은 25종의 영장류’ 목록 중에서 다음의 8종은 여전히 포함되어 있다:비단시파카, 델라쿠르랑구르, 흰머리랑구르, 회색정강이두크, 통킹들창코랑구르, 하이난검은볏긴팔원숭이, 크로스강고릴라 그리고 수마트라오랑우탄.[140] 아종 왈드론붉은콜로부스는 1993년부터 1999년까지 어느 곳에서도 흔적이 발견되지 않아, 최근 멸종된 것으로 선언되었다.[141] 몇몇 사냥꾼만이 발견하여 죽인 이래, 이 종이 발견될 전망은 거의 남아 있지 않다.[142]
함께 읽기
- 영장류 속 목록
- 영장류 종 목록
각주
↑ 가나다라마바사아 Groves, C.P. (2005). Wilson, D.E.; Reeder, D.M., 편집. 《Mammal Species of the World: A Taxonomic and Geographic Reference》 (영어) 3판. 존스 홉킨스 대학교 출판사. 111–184쪽. ISBN 0-801-88221-4. OCLC 62265494.
↑ 굿맨(Goodman, M.), 타글레(Tagle, D. A.), 피치(Fitch, D. H.), 베일리(Bailey, W.), 첼루스니악(Czelusniak, J.), 쿱(Koop, B. F.), 벤슨(Benson, P.) & 슬라이톰(Slightom, J. L.) (1990). “DNA 레벨 상의 영장류 진화 및 사람아과 분류”. 《분자진화학저널(Journal of Molecular Evolution)》 (영어) 30 (3): 260–266. PMID 2109087. doi:10.1007/BF02099995. CS1 관리 - 여러 이름 (링크)
↑ 가나다라마바 “영장류(브리태니커 백과사전 온라인)” (영어). 브리태니커 백과사전. 2008. 2008년 7월 21일에 확인함.
↑ 가나 채터지(Helen J Chatterjee), 호(Simon Y.W. Ho), 반즈(Ian Barnes) & 그로브스(Colin Groves) (2009). “초행렬 접근을 이용한 영장류 계통발생 및 분기 싯점 추산(Estimating the phylogeny and divergence times of primates using a supermatrix approach)”. 《BMC 진화생물학(BMC Evolutionary Biology)》 (영어) 9: 259. PMC 2774700. PMID 19860891. doi:10.1186/1471-2148-9-259. CS1 관리 - 여러 이름 (링크)
↑ Murphy W. J., E. Eizirik, W. E. Johnson, Y. P. Zhang, O. A. Ryder, S. J. O'Brien, 2001a. Molecular phylogenetics and the origins of placental mammals Nature 409:614-618. [1]
↑ Ulfur Arnason, et al. Mammalian mitogenomic relationships and the root of the eutherian tree. Proceedings of the National Academy of Science 99: 8151-8156. [2]
↑ Jan Ole Kriegs, Gennady Churakov, Jerzy Jurka, Jürgen Brosius, and Jürgen Schmitz (2007) Evolutionary history of 7SL RNA-derived SINEs in Supraprimates. Trends in Genetics 23 (4): 158-161 [3] (PDF version [4])
↑ 야넥카(Janečka, J. E.); 밀러(Miller, W.), 프링글(Pringle, T. H.), 위엔스(Wiens, F.), 지츠만(Zitzmann, A.), 헬젠(Helgen, K. M.), 스프링거(Springer, M. S.) & 머피(Murphy, W. J.) (2007). “Molecular and Genomic Data Identify the Closest Living Relative of Primates”. 《사이언스(Science)》 (영어) 318 (5851): 792–794. doi:10.1126/science.1147555. 2008년 8월 17일에 확인함. 더 이상 지원되지 않는 변수를 사용함 (도움말)
↑ Kavanagh, M. (1983). 《A Complete Guide to Monkeys, Apes and Other Primates》 (영어). New York: Viking Press. 18쪽. ISBN 0670435430.
↑ McKenna, M. C. and Bell, S. K. (1997). 《Classification of Mammals Above the Species Level》 (영어). New York: Columbia University Press. 329쪽. ISBN 023111012X. CS1 관리 - 여러 이름 (링크)
↑ Williams, B.A.; Kay, R.F.; Kirk, E.C. (2010). “유인원 기원에 관한 새로운 관점(New perspectives on anthropoid origins)”. 《미국 국립과학아카데미 회보(Proceedings of the National Academy of Sciences)》 (영어) 107 (11): 4797–4804. Bibcode:2010PNAS..107.4797W. PMC 2841917. PMID 20212104. doi:10.1073/pnas.0908320107.
↑ “Nova – Meet Your Ancestors” (영어). 미국 공영 방송망(PBS). 2008년 10월 24일에 확인함.
↑ “플레시아다피스(Plesiadapis)” (PDF) (영어). 노스다코타 지질조사국(North Dakota Geological Survey). 2008년 10월 24일에 확인함.
↑ Lee, M. (1999년 9월). “Molecular Clock Calibrations and Metazoan Divergence Dates”. 《Journal of Molecular Evolution》 (영어) 49 (3): 385–391. PMID 10473780. doi:10.1007/PL00006562. 더 이상 지원되지 않는 변수를 사용함 (도움말)
↑ “Scientists Push Back Primate Origins From 65 Million To 85 Million Years Ago” (영어). Science Daily. 2008년 10월 24일에 확인함.
↑ Tavaré, S., Marshall, C. R., Will, O., Soligo, C. & Martin R.D. (2002년 4월 18일). “Using the fossil record to estimate the age of the last common ancestor of extant primates”. 《네이처(Nature)》 (영어) 416 (6882): 726–729. PMID 11961552. doi:10.1038/416726a. CS1 관리 - 여러 이름 (링크) CS1 관리 - 날짜와 연도 (링크)
↑ Klonisch, T., Froehlich, C., Tetens, F., Fischer, B. & Hombach-Klonisch, S. (2001). “Molecular Remodeling of Members of the Relaxin Family During Primate Evolution”. 《Molecular Biology and Evolution》 (영어) 18 (3): 393–403. PMID 11230540. 2008년 8월 22일에 확인함. CS1 관리 - 여러 이름 (링크)
↑ 가나다라마 Horvath, J.; 외. (2008). “Development and Application of a Phylogenomic Toolkit: Resolving the Evolutionary History of Madagascar's Lemurs” (PDF). 《Genome Research》 (영어) 18 (3): 490. PMC 2259113. PMID 18245770. doi:10.1101/gr.7265208. 2008년 9월 10일에 원본 문서 (PDF)에서 보존된 문서. 2008년 8월 22일에 확인함. CS1 관리 - et al.의 직접적인 사용 (링크)
↑ 가나 Mittermeier, R., Ganzhorn, J., Konstant, W., Glander, K., Tattersall, I., Groves, C., Rylands, A., Hapke, A., Ratsimbazafy, J., Mayor, M., Louis, E., Rumpler, Y., Schwitzer, C. & Rasoloarison, R. (2008년 12월). “Lemur Diversity in Madagascar”. 《International Journal of Primatology》 (영어) 29 (6): 1607–1656. doi:10.1007/s10764-008-9317-y. 더 이상 지원되지 않는 변수를 사용함 (도움말) CS1 관리 - 여러 이름 (링크)
↑ 가나다라마 Sellers, Bill (2000년 10월 20일). “Primate Evolution” (PDF) (영어). University of Edinburgh. 13–17쪽. 2008년 10월 29일에 보존된 문서 (PDF). 2008년 10월 29일에 확인함.
↑ 가나다라 Hartwig, W. (2007). 〈Primate Evolution〉. Campbell, C., Fuentes, A., MacKinnon, K., Panger, M. & Bearder, S. 《Primates in Perspective》 (영어). Oxford University Press. 13–17쪽. ISBN 978-0-19-517133-4. CS1 관리 - 여러 이름 (링크)
↑ Franzen, Jens L.; et al. (2009). “Complete Primate Skeleton from the Middle Eocene of Messel in Germany: Morphology and Paleobiology”. 《PLoS ONE》 (영어) 4 (5): e5723. PMC 2683573. PMID 19492084. doi:10.1371/journal.pone.0005723. 더 이상 지원되지 않는 변수를 사용함 (도움말)
↑ Ciochon, R. & Fleagle, J. (1987). 《Primate Evolution and Human Origins》 (영어). Menlo Park, California: Benjamin/Cummings. 72쪽. ISBN 9780202011752. CS1 관리 - 여러 이름 (링크)
↑ 가나 Garbutt, N. (2007). 《Mammals of Madagascar, A Complete Guide》 (영어). A&C Black Publishers. 85–86쪽. ISBN 978-0-300-12550-4.
↑ 가나 Mittermeier, R.A.; et al. (2006). 《Lemurs of Madagascar》 (영어) 2판. Conservation International. 23–26쪽. ISBN 1-881173-88-7. 더 이상 지원되지 않는 변수를 사용함 (도움말)
↑ Shekelle, M. (2005). “Evolutionary Biology of Tarsiers” (영어). 2008년 8월 22일에 확인함.
↑ Schmidt, T.; 외. (2005). “Rapid electrostatic evolution at the binding site for cytochrome c on cytochrome c oxidase in anthropoid primates”. 《Proceedings of the National Academy of Sciences of the United States of America》 (영어) 102 (18): 6379–6384. PMC 1088365. PMID 15851671. doi:10.1073/pnas.0409714102. 2008년 8월 22일에 확인함. CS1 관리 - et al.의 직접적인 사용 (링크)
↑ Marivaux, L.; 외. (2005년 6월 14일). “Anthropoid primates from the Oligocene of Pakistan (Bugti Hills): Data on early anthropoid evolution and biogeography”. 《Proceedings of the National Academy of Sciences of the United States of America》 (영어) 102 (24): 8436–8441. PMC 1150860. PMID 15937103. doi:10.1073/pnas.0503469102. 2008년 8월 22일에 확인함. CS1 관리 - et al.의 직접적인 사용 (링크)
↑ Schrago, C.G. & Russo, C.A.M. (2003). “Timing the Origin of New World Monkeys” (PDF Reprint). 《Molecular Biology and Evolution》 (영어) 20 (10): 1620–1625. PMID 12832653. doi:10.1093/molbev/msg172. CS1 관리 - 여러 이름 (링크)
↑ Houle, A. (1999). “The origin of platyrrhines: An evaluation of the Antarctic scenario and the floating island model”. 《American Journal of Physical Anthropology》 (영어) 109 (4): 541–559. PMID 10423268. doi:10.1002/(SICI)1096-8644(199908)109:4<541::AID-AJPA9>3.0.CO;2-N.
↑ Andrews, P. & Kelley, J. (2007). “Middle Miocene Dispersals of Apes”. 《Folia Primatologica》 (영어) 78 (5-6): 328–343. PMID 17855786. doi:10.1159/000105148. CS1 관리 - 여러 이름 (링크)
↑ 가나다라마바사아자차카타파하거너더러머버 Strier, K. (2007). 《Primate Behavioral Ecology》 (영어) 3판. Allyn & Bacon. 7, 64, 71, 77, 182–185, 273–280, 284, 287–298쪽. ISBN 0-205-44432-6. CS1 관리 - 추가 문구 (링크)
↑ Pough, F. W., Janis, C. M. & Heiser, J. B. (2005). 〈Primate Evolution and the Emergence of Humans〉. 《Vertebrate Life》 (영어) 7판. Pearson. 650쪽. ISBN 0-13-127836-3. 지원되지 않는 변수 무시됨:|초판출판년도=(도움말) CS1 관리 - 여러 이름 (링크) CS1 관리 - 추가 문구 (링크)
↑ Groves, C. P. 《Primate Taxonomy》 (영어). Smithsonian Institution Press. ISBN 1-56098-872-X. 지원되지 않는 변수 무시됨:|출판 년도=(도움말)
↑ Rylands, A. B. & Mittermeier, R. A. (2009). 〈The Diversity of the New World Primates (Platyrrhini)〉. Garber, P. A., Estrada, A., Bicca-Marques, J. C., Heymann, E. W. & Strier, K. B. 《South American Primates: Comparative Perspectives in the Study of Bahavior, Ecology, and Conservation》 (영어). Springer. ISBN 978-0-387-78704-6. CS1 관리 - 여러 이름 (링크)
↑ 칼 폰 린네 (1758). 《Sistema naturae per regna tria Naturae, secundum classes, ordines, genera, species, cum characteribus differentiis, synonimis locis. Tomus I》 (영어). Impensis direct. Laurentii Salvii, Holmia. 20–32쪽.
↑ 칼 폰 린네 (1735). 《Sistema naturae sive regna tria Naturae systematice proposita per classes, ordines, genera, & species》 (영어). apud Theodorum Haak, Lugduni Batavorum. s.p.쪽.
↑ Blainville, H. (1839). 〈Nouvelle classification des Mammifères〉. 《Annales Françaises et Etrangères d'Anatomie et de Physiologie Appliquées à la Médicine et à l'Histoire Naturelle, 3》 (영어). 268–269쪽.
↑ McKenna, M. C. & Bell, S. K. (1997). 《Classification of Mammals: Above the species level》 (영어). New York: Columbia University Press. 631쪽. ISBN 0-231-11013-8. CS1 관리 - 여러 이름 (링크)
↑ Strier, K. (2007). 《Primate Behavioral Ecology》 (영어) 3판. 50–53쪽. ISBN 0-295-44432-6|isbn=값 확인 필요: checksum (도움말). CS1 관리 - 추가 문구 (링크)
↑ Tenaza, R. (1984). “Songs of hybrid gibbons (Hylobates lar × H. muelleri)”. 《American Journal of Primatology》 (영어) 8 (3): 249–253. doi:10.1002/ajp.1350080307.
↑ 가나 Bernsteil, I. S. (1966). “Naturally occurring primate hybrid”. 《사이언스(Science)》 (영어) 154 (3756): 1559–1560. PMID 4958933. doi:10.1126/science.154.3756.1559.
↑ Sugawara, K. (1979). “Sociological study of a wild group of hybrid baboons between Papio anubis and P. hamadryas in the Awash Valley, Ethiopia”. 《Primates》 (영어) 20 (1): 21–56. doi:10.1007/BF02373827.
↑ Jolly, C. J.; 외. (1997). “Intergeneric Hybrid Baboons”. 《International Journal of Primatology》 (영어) 18 (4): 597–627. doi:10.1023/A:1026367307470. CS1 관리 - et al.의 직접적인 사용 (링크)
↑ 가나다라마바사아자차카타파하거너더 Pough, F. W., Janis, C. M. & Heiser, J. B. (2005) [1979]. 〈Characteristics of Primates〉. 《Vertebrate Life》 (영어) 제7판. Pearson. 630쪽. ISBN 0-13-27836-3|isbn=값 확인 필요: length (도움말). CS1 관리 - 여러 이름 (링크) CS1 관리 - 추가 문구 (링크)
↑ 가나 Soligo, C., Müller, A.E. (1999). “Nails and claws in primate evolution”. 《Journal of Human Evolution》 (영어) 36: 97–114. doi:10.1006/jhev.1998.0263. CS1 관리 - 여러 이름 (링크)
↑ 가나다라마바사아자차카타파 Macdonald, David (2006). 《The Encyclopedia of Mammals》 (영어). The Brown Reference Group plc. 290–307쪽. ISBN 0-681-45659-0. 필요 이상의 변수가 사용됨:|저자=및|성=(도움말)
↑ 가나다 White, T. & Kazlev, A. (2006년 1월 8일). “Archonta: Primates” (영어). Palaeos. 2008년 5월 12일에 원본 문서에서 보존된 문서. 2008년 6월 3일에 확인함. CS1 관리 - 여러 이름 (링크)
↑ 가나 Pough, F. W., Janis, C. M. & Heiser, J. B. (2005) [1979]. 〈Primate Societies〉. 《Vertebrate Life》 (영어) 7판. Pearson. 621–623쪽. ISBN 0-13-127836-3. CS1 관리 - 여러 이름 (링크) CS1 관리 - 추가 문구 (링크)
↑ Campbell, B. G. & Loy, J. D. (2000). 《Humankind Emerging (8th edition)》 (영어). Allyn & Bacon. 85쪽. ISBN 0673523640. CS1 관리 - 여러 이름 (링크)
↑ 가나 Aiello, L. & Dean, C. (1990). 《An Introduction to Human Evolutionary Anatomy》 (영어). Academic Press. 193쪽. ISBN 0120455900. CS1 관리 - 여러 이름 (링크)
↑ 가나다라마바사 Myers, P. (1999). “"Primates" (On-line)” (영어). Animal Diversity Web. 2008년 6월 3일에 확인함.
↑ Ash, M. M., Nelson, S. J. & Wheeler, R. C. (2003). 《Wheeler's Dental Anatomy, Physiology, and Occlusion》 (영어). W.B. Saunders. 12쪽. ISBN 9780721693828. CS1 관리 - 여러 이름 (링크)
↑ Arrese, C. A.; 외. (2005). “Cone topography and spectral sensitivity in two potentially trichromatic marsupials, the quokka (Setonix brachyurus) and quenda (Isoodon obesulus)”. 《Proceedings of Biological Science》 (영어) 272 (1565): 791–796. PMC 1599861. PMID 15888411. doi:10.1098/rspb.2004.3009. CS1 관리 - et al.의 직접적인 사용 (링크)
↑ Bowmaker, J. K. & Astell, S. (1991). “Photosensitive and photostable pigments in the retinae of Old World monkeys” (pdf). 《Journal of Experimental Biology》 (영어) 156: 1–19. ISSN 0022-0949. PMID 2051127. 2008년 6월 16일에 확인함. CS1 관리 - 여러 이름 (링크)
↑ Surridge, A. K., and D. Osorio (2003). “Evolution and selection of trichromatic vision in primates”. 《Trends in Ecology and Evolution》 (영어) 18: 198–205. doi:10.1016/S0169-5347(03)00012-0. CS1 관리 - 여러 이름 (링크)
↑ Lucas, P. W. & Dominy, N. J. (2003). “Evolution and function of routine trichromatic vision in primates”. 《Evolution》 (영어) 57 (11): 2636–2643. PMID 14686538. doi:10.1554/03-168. CS1 관리 - 여러 이름 (링크)
↑ 가나다라 Sussman, R. W. (2003). 《Primate Ecology and Social Structure, Volume 2: New World Monkeys》 (영어) 개정 초판. Needham Heights, MA: Pearson Custom Publishing & Prentice Hall. 77–80, 132–133, 141–143쪽. ISBN 0-536-74364-9. CS1 관리 - 추가 문구 (링크)
↑ Surridge, A. K. & Osorio, D. (2003). “Evolution and selection of trichromatic vision in primates”. 《Trends in Ecology and Evolution》 (영어) 18: 198–205. doi:10.1016/S0169-5347(03)00012-0. CS1 관리 - 여러 이름 (링크)
↑ Ralls, K. (1976). “Mammals in Which Females are Larger Than Males”. 《The Quarterly Review of Biology》 (영어) 51 (2): 245. PMID 785524. doi:10.1086/409310.
↑ Lindstedtand & Boyce (1985). “Seasonality, Fasting Endurance, and Body Size in Mammals”. 《The American Naturalist》 (영어) 125: 873. doi:10.1086/284385.
↑ Frisch, J. E. (1963). “Sex-differences in the canines of the gibbon (Hylobates lar)”. 《Primates》 (영어) 4 (2): 1. doi:10.1007/BF01659148.
↑ Kay, R. F. (1975). “The functional adaptations of primate molar teeth”. 《American Journal of Physical Anthropology》 (영어) 43 (2): 195–215. PMID 810034. doi:10.1002/ajpa.1330430207.
↑ Crook, J. H. (1972). 〈Sexual selection, dimorphism, and social organization in the primates〉. Campbell, B. G. 《Sexual selection and the descent of man》 (영어). Aldine Transaction. 246쪽. ISBN 978-0202020051.
↑ 가나 Cheverud, J. M., Dow, M. M. & Leutenegger, W. (1985). “The quantitative assessment of phylogenetic constraints in comparative analyses: Sexual dimorphism in body weight among primates”. 《Evolution》 (영어) 39 (6): 1335–1351. doi:10.2307/2408790. CS1 관리 - 여러 이름 (링크)
↑ Leutenegger, W. & Cheverud, J. M. (1982). “Correlates of sexual dimorphism in primates: Ecological and size variables”. 《International Journal of Primatology》 (영어) 3 (4): 387. doi:10.1007/BF02693740. CS1 관리 - 여러 이름 (링크)
↑ 가나 Plavcan, J. M. (2001). “Sexual dimorphism in primate evolution”. 《American Journal of Physical Anthropology》 (영어) 33: 25–53. PMID 11786990. doi:10.1002/ajpa.10011.
↑ O'Higgins, P. & Collard, M. (2002). “Sexual dimorphism and facial growth in papionine monkeys”. 《동물학 저널(Journal of Zoology)》 (영어) 257 (2): 255–272. doi:10.1017/S0952836902000857. CS1 관리 - 여러 이름 (링크)
↑ Sussman, R. W. (1999). 《영장류 생태 및 사회 구조 1권: 로리스, 여우원숭이 및 안경원숭이》 (영어). Needham Heights, MA: Pearson Custom Publishing & Prentice Hall. 78, 89–90, 108, 121–123, 233쪽. ISBN 0-536-02256-9.
↑ Wrangham, R. W. (1982). 〈Mutualism, kinship and social evolution〉. 《Current Problems in Sociobiology》 (영어). 캠브리지 대학교 출판부. 269–289쪽. ISBN 0521242037.
↑ Fiore, A. D. & Campbell, C. J. (2007). 〈The Atelines〉. Campbell, C. J., Fuentes, A., MacKinnon, K. C., Panger, M. & Bearder, S. K. 《Primates in Perspective》 (영어). 옥스포드 대학교 출판부. 175쪽. ISBN 978-0-19-517133-4. CS1 관리 - 여러 이름 (링크)
↑ Bartlett, T. Q. (2007). 〈The Hylobatidae〉. Campbell, C. J., Fuentes, A., MacKinnon, K. C., Panger, M. & Bearder, S. K. 《Primates in Perspective》 (영어). Oxford University Press. 283쪽. ISBN 978-0-19-517133-4. CS1 관리 - 여러 이름 (링크)
↑ Knott, C. D. & Kahlenberg, S. M. (2007). 〈Orangutans in Perspective〉. Campbell, C. J., Fuentes, A., MacKinnon, K. C., Panger, M. & Bearder, S. K. 《Primates in Perspective》 (영어). Oxford University Press. 294쪽. ISBN 978-0-19-517133-4. CS1 관리 - 여러 이름 (링크)
↑ Constable, J. L.; 외. (2001). “Noninvasive paternity assignment in Gombe chimpanzees”. 《Molecular Ecology》 (영어) 10 (5): 1279–1300. PMID 11380884. doi:10.1046/j.1365-294X.2001.01262.x. CS1 관리 - et al.의 직접적인 사용 (링크)
↑ 가나다라마 Rowe, N. (1996). 《The Pictorial Guide to the Living Primates》 (영어). Pogonias Press. 4, 139, 143, 154, 185, 223쪽. ISBN 0-9648825-0-7.
↑ Pough, F. W., Janis, C. M. & Heiser, J. B. (2005). 〈Primate Societies〉. 《Vertebrate Life》 (영어) 7판. Pearson. 621–623쪽. ISBN 0-13-127836-3. 지원되지 않는 변수 무시됨:|초판년도=(도움말) CS1 관리 - 여러 이름 (링크) CS1 관리 - 추가 문구 (링크)
↑ Smuts, B.B., Cheney, D.L. Seyfarth, R.M., Wrangham, R.W., & Struhsaker, T.T. (Eds.) (1987). Primate Societies. Chicago: University of Chicago Press for articles on the structure and function of various primate societies.
↑ Shultz, S. & Thomsett, S. (2007). 〈Interactions between African Crowned Eagles and Their Prey Community〉. McGraw, W., Zuberbuhler, K. & Noe, R. 《Monkeys of Tai Forest, An African Primate Community》 (영어). Cambridge University Press. 181쪽. ISBN 0-521-81633-5. CS1 관리 - 여러 이름 (링크)
↑ Bshary, R. (2007). 〈Interactions between Red Colobus Monkeys and Chimpanzees〉. McGraw, W., Zuberbuhler, K. & Noe, R. 《=Monkeys of Tai Forest, An African Primate Community》 (영어). Cambridge University Press. 155–170쪽. ISBN 0-521-81633-5. CS1 관리 - 여러 이름 (링크)
↑ Stanford, C. (1998). 《Chimpanzee and Red Colobus : the ecology of predator and prey》 (영어). Harvard University Press. 130–138, 233쪽. ISBN 0-674-00722-0.
↑ 가나 Boinski, S. (2000). 〈Social Manipulation Within and Between Troops Mediates Primate Group Movement〉. Boinski, S. and Garber, P. 《On the Move : how and why animals travel in groups》 (영어). University of Chicago Press. 447–448쪽. ISBN 0-226-06340-2. CS1 관리 - 여러 이름 (링크)
↑ Boesch, C. & Boesch, H. (1990). “Tool Use and Tool Making in Wild Chimpanzees”. 《Folia Primatol》 (영어) 54 (1-2): 86–99. PMID 2157651. doi:10.1159/000156428. CS1 관리 - 여러 이름 (링크)
↑ Westergaard, G. C.; 외. (1998). “Why some capuchin monkeys (Cebus apella) use probing tools (and others do not)”. 《Journal of Comparative Psychology》 (영어) 112 (2): 207–211. PMID 9642788. doi:10.1037/0735-7036.112.2.207. CS1 관리 - et al.의 직접적인 사용 (링크)
↑ de Waal, F. B. M. & Davis, J. M. (2003). “Capuchin cognitive ecology: cooperation based on projected returns”. 《Neuropsychologia》 (영어) 41 (2): 221–228. PMID 12459220. doi:10.1016/S0028-3932(02)00152-5. CS1 관리 - 여러 이름 (링크)
↑ Paar, L. A., Winslow, J. T., Hopkins, W. D. & de Waal, F. B. M. (2000). “Recognizing facial cues: Individual discrimination by chimpanzees (Pan troglodytes) and rhesus monkeys (Macaca mulatta)”. 《Journal of Comparative Psychology》 (영어) 114 (1): 47–60. PMC 2018744. PMID 10739311. doi:10.1037/0735-7036.114.1.47. CS1 관리 - 여러 이름 (링크)
↑ Paar, L. A. & de Waal, F. B. M. (1999). “Visual kin recognition in chimpanzees”. 《Nature》 (영어) 399 (6737): 647. PMID 10385114. doi:10.1038/21345. CS1 관리 - 여러 이름 (링크)
↑ Fujita, K., Watanabe, K., Widarto, T. H. & Suryobroto, B. (1997). “Discrimination of macaques by macaques: The case of sulawesi species”. 《Primates》 (영어) 38 (3): 233–245. doi:10.1007/BF02381612. CS1 관리 - 여러 이름 (링크)
↑ Call, J. (2001). “Object permanence in orangutans (Pongo pygmaeus), chimpanzees (Pan troglodytes), and children (Homo sapiens)”. 《Journal of Comparative Psychology》 (영어) 115 (2): 159–171. PMID 11459163. doi:10.1037/0735-7036.115.2.159.
↑ Itakura, S. & Tanaka, M. (1998). “Use of experimenter-given cues during object-choice tasks by chimpanzees (Pan troglodytes), an orangutan (Pongo pygmaeus), and human infants (Homo sapiens)”. 《Journal of Comparative Psychology》 (영어) 112 (2): 119–126. PMID 9642782. doi:10.1037/0735-7036.112.2.119. CS1 관리 - 여러 이름 (링크)
↑ Gouteux, S., Thinus-Blanc, C. & Vauclair, J. (2001). “Rhesus monkeys use geometric and nongeometric information during a reorientation task”. 《Journal of Experimental Psychology: General》 (영어) 130 (3): 505–519. doi:10.1037/0096-3445.130.3.505. CS1 관리 - 여러 이름 (링크)
↑ Tomasello, M. & Call, J. (1997). 《Primate Cognition》 (영어). Oxford University Press US. ISBN 9780195106244. CS1 관리 - 여러 이름 (링크)
↑ Liman, E. R. & Innan, H. (2003). “Relaxed selective pressure on an essential component of pheromone transduction in primate evolution” (pdf). 《Proceedings of the National Academy of Sciences of the United States of America》 (영어) 100 (6): 3328–3332. PMC 152292. PMID 12631698. doi:10.1073/pnas.0636123100. 2008년 7월 23일에 확인함. CS1 관리 - 여러 이름 (링크)
↑ Egnor, R., Miller, C. & Hauser, M.D. (2004). 〈Nonhuman Primate Communication〉 (PDF). 《Encyclopedia of Language and Linguistics》 (pdf)|format=은|url=을 필요로 함 (도움말) (영어) 2판. Elsevier. ISBN 0080442994. 2008년 7월 23일에 확인함. CS1 관리 - 여러 이름 (링크) CS1 관리 - 추가 문구 (링크)
↑ Milton, C. (August 1993) "Diet and Primate Evolution" Scientific American vol.269 pp.70–77
↑ Milliken, G. W., Ward, J. P. & Erickson, C. J. (1991). “Independent digit control in foraging by the aye-aye (Daubentonia madagascariensis)”. 《Folia Primatologica》 (영어) 56 (4): 219–224. doi:10.1159/000156551. CS1 관리 - 여러 이름 (링크)
↑ Hiller, C. (2000). “Theropithecus gelada”. 《Animal Diversity Web》 (영어). 2008년 8월 8일에 확인함.
↑ Wright, P., Simmons, E. & Gursky, S. (2003). 〈Introduction〉. Wright, P., Simmons, E. & Gursky, S. 《Tarsiers Past, Present and Future》 (영어). Rutgers University Press. 1쪽. ISBN 0-8135-3236-1. CS1 관리 - 여러 이름 (링크)
↑ Bshary, R. (2007). 〈Interactions between Red Colobus Monkeys and Chimpanzees〉. McGraw, W., Zuberbuhler, K. & Noe, R. 《Monkeys of the Taï Forest : an African primate community》 (영어). Cambridge University Press. 155–170쪽. ISBN 0-521-81633-5. CS1 관리 - 여러 이름 (링크)
↑ Stanford, C. (1998). 《Chimpanzee and red colobus : the ecology of predator and prey》 (영어). Harvard University Press. 130–138, 233쪽. ISBN 0-674-00722-0.
↑ Reed, K. & Fleagle, J. (1995년 8월 15일). “Geographic and climatic control of primate diversity” (PDF). 《Proceedings of the National Academy of Sciences of the United States of America》 (영어) 92 (17): 7874–7876. PMC 41248. PMID 7644506. doi:10.1073/pnas.92.17.7874. CS1 관리 - 여러 이름 (링크) CS1 관리 - 날짜와 연도 (링크)
↑ Chapman, C. & Russo, S. (2007). 〈Primate Seed Dispersal〉. Campbell, C. J., Fuentes, A., MacKinnon, K. C., Panger, M. & Bearder, S. K. 《Primates in Perspective》 (영어). Oxford University Press. 510쪽. ISBN 978-0-19-517133-4. CS1 관리 - 여러 이름 (링크)
↑ Long, Y. C., Kirkpatrick, R. C., Zhong, T., and Xiao, L. (1994). “Report on the distribution, population, and ecology of the Yunnan snub-nosed monkey (Rhinopithecus bieti)”. 《Primates》 (영어) 35: 241–250. doi:10.1007/BF02382060. CS1 관리 - 여러 이름 (링크)
↑ Schaller, G. B. (1963). 《The Mountain Gorilla: Ecology and Behavior》 (영어). Chicago: University Chicago Press. ISBN 978-0226736358.
↑ Wolfe, L. D. & Fuentes, A. (2007). 〈Ethnoprimatology〉. Campbell, C. J., Fuentes, A., MacKinnon, K. C., Panger, M. & Bearder, S. K. 《Primates in Perspective》 (영어). Oxford University Press. 692쪽. ISBN 978-0-19-517133-4. CS1 관리 - 여러 이름 (링크)
↑ Veterinary Clinics of North America, Small Animal Practice (1987). “Zoonoses Acquired from Pet Primates” (영어) 17 (1): 219–240. 2008년 8월 11일에 확인함.
↑ 가나 Cavalieri, P. & Singer, P. “Declaration on Great Apes” (영어). Great Ape Project. 2008년 8월 20일에 원본 문서에서 보존된 문서. 2008년 6월 16일에 확인함. CS1 관리 - 여러 이름 (링크)
↑ Glendinning, L. (2008년 6월 26일). “Spanish parliament approves 'human rights' for apes” (영어). The Guardian. 2008년 11월 10일에 확인함.
↑ Singer, P. (2008년 7월 18일). “Of great apes and men” (영어). The Guardian. 2008년 11월 10일에 확인함.
↑ Mager, B. (2006년 7월 6일). “The HSUS Applauds the U.S. Senate for Unanimously Passing the Captive Primate Safety Act” (영어). 미국 휴메인 소사이어티. 2008년 12월 11일에 원본 문서에서 보존된 문서. 2008년 11월 10일에 확인함.
↑ 가나 Workman, C. (2004). “Primate conservation in Vietnam: toward a holistic environmental narrative”. 《American Anthropologist》 (영어) 106 (2): 346–352. doi:10.1525/aa.2004.106.2.346.
↑ “IPPL News: The US Pet Monkey Trade” (영어). 국제영장류보호연맹. 2003. 2008년 7월 25일에 원본 문서에서 보존된 문서. 2008년 8월 4일에 확인함.
↑ Bushnell, D. (1958). “The Beginnings of Research in Space Biology at the Air Force Missile Development Center, 1946-1952”. 《History of Research in Space Biology and Biodynamics》 (영어). 미국 항공우주국. 2008년 8월 18일에 확인함.
↑ Blumenthal, D. (1987년 6월 17일). “Monkeys as Helpers To Quadriplegics At Home”. 《The New York Times》 (영어). 2008년 10월 8일에 확인함. CS1 관리 - 날짜와 연도 (링크)
↑ 가나 “The supply and use of primates in the EU” (영어). European Biomedical Research Association. 1996. 2012년 1월 17일에 원본 문서에서 보존된 문서. 2008년 8월 18일에 확인함.
↑ Chen, F. C. & Li, W. H. (2001). “Genomic divergences between humans and other hominoids and the effective population size of the common ancestor of humans and chimpanzees”. 《American Journal of Human Genetics》 (영어) 68 (2): 444–456. PMC 1235277. PMID 11170892. doi:10.1086/318206. 2008년 10월 8일에 확인함. CS1 관리 - 여러 이름 (링크)
↑ Conlee, K. M., Hoffeld, E. H. & Stephens, M. L. (2004). “A Demographic Analysis of Primate Research in the United States” (PDF). 《ATLA (Alternatives to Laboratory Animals)》 (영어) 32 (Sup 1): 315–322. 2008년 6월 25일에 원본 문서 (pdf)에서 보존된 문서. 2008년 10월 8일에 확인함. CS1 관리 - 여러 이름 (링크)
↑ presented to Parliament by the Secretary of State for the Home Department by Command of Her Majesty, July 2006. (2006년 7월). 《Statistics of scientific procedures on living animals: Great Britain 2005》 (pdf) (영어). The Stationery Office. ISBN 0101687729. 2008년 6월 16일에 확인함. 더 이상 지원되지 않는 변수를 사용함 (도움말)
↑ “Nonhuman Primates: Research Animals”. 《Animal Welfare Information Center》 (영어). United States Department of Agriculture. 2008년 10월 3일에 원본 문서에서 보존된 문서. 2008년 7월 14일에 확인함.
↑ “Directive 86/609” (영어). European Coalition to End Animal Experiments. 2008년 9월 29일에 원본 문서에서 보존된 문서. 2008년 10월 8일에 확인함.
↑ Grubb, P.; 외. (1998). 〈The Sierra Leone monkey drives〉. 《Mammals of Ghana, Sierra Leone, and the Gambia》 (영어). St. Ives: Trendrine. 214–219쪽. ISBN 0951256246. CS1 관리 - et al.의 직접적인 사용 (링크)
↑ 가나 Chapman, C. A. & Peres, C. A. (2001). “Primate conservation in the new millennium: the role of scientists”. 《Evolutionary Anthropology》 (영어) 10: 16–33. doi:10.1002/1520-6505(2001)10:1<16::AID-EVAN1010>3.0.CO;2-O. CS1 관리 - 여러 이름 (링크)
↑ 가나 Mittermeier, R. A. & Cheney, D. L. (1987). 〈Conservation of primates and their habitats〉. Smuts, B. B., Cheney, D. L., Seyfarth, R. M., Wrangham, R. W. & Struhsaker, T. T. 《Primate Societies》 (영어). Chicago: University of Chicago Press. 477–490쪽. CS1 관리 - 여러 이름 (링크)
↑ 가나 Southwick, C. H. & Siddiqi, M. F. (2001). “Status, conservation and management of primates in India” (PDF). 《Envis Bulletin: Wildlife and Protected Areas》 (영어) 1 (1): 81–91. 2008년 10월 1일에 원본 문서 (pdf)에서 보존된 문서. 2008년 8월 4일에 확인함. CS1 관리 - 여러 이름 (링크)
↑ 가나다라 Cowlishaw, G. & Dunbar, R. (2000). 《Primate Conservation Biology》 (영어). Chicago: University of Chicago Press. ISBN 9780226116372. CS1 관리 - 여러 이름 (링크)
↑ Van Schaik, C. P., Monk, K. A. & Robertson, J. M. Y. (2001). “Dramatic decline in orangutan numbers in the Leuser Ecosystem, northern Sumatra”. 《Oryx》 (영어) 35 (1): 14–25. doi:10.1046/j.1365-3008.2001.00150.x. CS1 관리 - 여러 이름 (링크)
↑ Purvis, A., Gittleman, J. L., Cowlishaw, G. & Mace, G. M. (2000). “Predicting extinction risk in declining species”. 《Proceedings of the Royal Society of London》 (영어) 267 (1456): 1947–1952. PMC 1690772. PMID 11075706. doi:10.1098/rspb.2000.1234. CS1 관리 - 여러 이름 (링크)
↑ 가나다 Fa, J. E., Juste, J., Perez de Val, J. & Castroviejo, J. (1995). “Impact of market hunting on mammal species in Equatorial Guinea”. 《Conservation Biology》 (영어) 9 (5): 1107–1115. doi:10.1046/j.1523-1739.1995.9051107.x. CS1 관리 - 여러 이름 (링크)
↑ Hill, C. M. (1997). “Crop-raiding by wild vertebrates: The farmer's perspective in an agricultural community in western Uganda”. 《International Journal of Pest Management》 (영어) 43 (1): 77–84. doi:10.1080/096708797229022.
↑ Hill, C. M. (2002). “Primate conservation and local communities: Ethical issues and debates”. 《American Anthropologist》 (영어) 104 (4): 1184–1194. doi:10.1525/aa.2002.104.4.1184.
↑ 〈Primate〉. 《Encyclopædia Britannica Online》. Encyclopædia Britannica, Inc. 2008. 2008년 7월 21일에 확인함.
↑ Choudhury, A. (2001). “Primates in Northeast India: an overview of their distribution and conservation status” (PDF). 《Envis Bulletin: Wildlife and Protected Areas》 (영어) 1 (1): 92–101. 2008년 10월 1일에 원본 문서 (pdf)에서 보존된 문서. 2008년 8월 4일에 확인함.
↑ Kumara, H. N. & Singh, M. (2004). “Distribution and abundance of primates in rainforests of the Western Ghats, Karnataka, India and the conservation of Macaca silenus”. 《International Journal of Primatology》 (영어) 25 (5): 1001–1018. doi:10.1023/B:IJOP.0000043348.06255.7f. CS1 관리 - 여러 이름 (링크)
↑ Nijman, V. (2004). “Conservation of the Javan gibbon Hylobates moloch: population estimates, local extinction, and conservation priorities” (pdf). 《The Raffles Bulletin of Zoology》 (영어) 52 (1): 271–280. 2008년 8월 4일에 확인함.
↑ O'Brien, T. G., Kinnaird, M. F., Nurcahyo, A., Iqbal, M. & Rusmanto, M. (2004). “Abundance and distribution of sympatric gibbons in a threatened Sumatran rain forest”. 《International Journal of Primatology》 (영어) 25 (2): 267–284. doi:10.1023/B:IJOP.0000019152.83883.1c. CS1 관리 - 여러 이름 (링크)
↑ Estrada, A., Coates-Estrada, R. & Meritt, D. (1994). “Non-flying mammals and landscape changes in the tropical forest region of Los Tuxtlas, Mexico”. 《Ecography》 (영어) 17: 229–241. doi:10.1111/j.1600-0587.1994.tb00098.x. CS1 관리 - 여러 이름 (링크)
↑ Marsh, L. K. (2003). 〈The nature of fragmentation.〉. Marsh, L. K. 《Primates in Fragments: Ecology and Conservation》. New York: Kluwer Academic/Plenum Publishers. 1–10쪽.
↑ Turner, I. M. (1996). “Species loss in fragments of tropical rain forest: a review of the evidence”. 《Journal of Applied Ecology》 (영어) 33 (2): 200–209. doi:10.2307/2404743.
↑ Chiarello, A.G. (2003). 〈Primates of the Brazilian Atlantic forest: the influence of forest fragmentation on survival〉. Marsh, L. K. 《Primates in Fragments: Ecology and Conservation》. New York: Kluwer Academic/Plenum Publishers. 99–121쪽. ISBN 9780306476969.
↑ Pope, T.R. (1996). 〈Socioecology, population fragmentation, and patterns of genetic loss in endangered primates〉. Avise, J. & Hamrick, J. 《Conservation Genetics: Case Histories from Nature》 (영어). Norwell: Kluwer Academic Publishers. 119–159쪽. ISBN 9780412055812. CS1 관리 - 여러 이름 (링크)
↑ Mittermeier, R. A.; 외. (2007). “Primates in Peril: The World's 25 Most Endangered Primates, 2006 – 2008” (PDF). 《Primate Conservation》 (영어) 22 (22): 1–40. doi:10.1896/052.022.0101. 2008년 7월 20일에 원본 문서 (pdf)에서 보존된 문서. 2008년 8월 4일에 확인함. CS1 관리 - et al.의 직접적인 사용 (링크)
↑ Oates, J. F., Abedi-Lartey, M., McGraw, W. S., Struhsaker, T. T. & Whitesides, G. H. (2000). “Extinction of a West African Red Colobus Monkey”. 《The Journal of the Society for Conservation Biology》 (영어) 14 (5): 1526. doi:10.1046/j.1523-1739.2000.99230.x. CS1 관리 - 여러 이름 (링크)
↑ McGraw, W. S. (2005). “Update on the Search for Miss Waldron’s Red Colobus Monkey”. 《International Journal of Primatology》 (영어) 26 (3): 605–619. doi:10.1007/s10764-005-4368-9.
외부 링크
| 위키미디어 공용에 관련된 미디어 자료가 있습니다. 영장류 |
| 위키생물종에 관련된 분류 및 자료가 있습니다. 영장류 |
(영어) 영장류 정보넷(Primate Info Net)
(영어) 동물다양성웹(Animal Diversity Web)의 영장류(Primates) 정보
(영어) 교토 대학의 영장류 연구소
(영어) 고해상도 세포구축학적 영장류 뇌 사진/도해(High-Resolution Cytoarchitectural Primate Brain Atlases)
(영어) 유러피안 영장류 네트워크(EUPRIM-Net: European Primate Network)
(영어) 영장류 사진: 자연사 콜렉션(PrimateImages: Natural History Collection)
(영어) 생물의 나무(Tree of Life) 웹프로젝트(web project)의 영장류
분류:
- 영장류
- 칼 폰 린네가 명명한 분류군
(window.RLQ=window.RLQ||[]).push(function()mw.config.set("wgPageParseReport":"limitreport":"cputime":"1.552","walltime":"1.678","ppvisitednodes":"value":9120,"limit":1000000,"ppgeneratednodes":"value":0,"limit":1500000,"postexpandincludesize":"value":402258,"limit":2097152,"templateargumentsize":"value":14928,"limit":2097152,"expansiondepth":"value":16,"limit":40,"expensivefunctioncount":"value":0,"limit":500,"unstrip-depth":"value":0,"limit":20,"unstrip-size":"value":207470,"limit":5000000,"entityaccesscount":"value":1,"limit":400,"timingprofile":["100.00% 1306.122 1 -total"," 72.36% 945.165 1 틀:각주"," 23.84% 311.361 68 틀:저널_인용"," 21.79% 284.647 50 틀:서적_인용"," 6.38% 83.315 18 틀:웹_인용"," 6.17% 80.600 8 틀:둘러보기_상자/중첩"," 5.60% 73.176 2 틀:둘러보기_상자"," 4.91% 64.149 1 틀:MSW3_Groves"," 3.25% 42.497 1 틀:포유류"," 2.88% 37.616 1 틀:영장류"],"scribunto":"limitreport-timeusage":"value":"0.689","limit":"10.000","limitreport-memusage":"value":3789089,"limit":52428800,"cachereport":"origin":"mw1228","timestamp":"20190406144823","ttl":2592000,"transientcontent":false);mw.config.set("wgBackendResponseTime":164,"wgHostname":"mw1266"););

